Learn more about the Reproductive System
In this module, you will learn more about
- Male anatomy and physiology
- Female anatomy and physiology
- Cell division: mitosis and meiosis
- Spermatogenesis & Oogenesis
- The ovarian and uterine cycle
- Pregnancy, development and birth
- Inheritance
Learn even more: See Cells, Chemistry, Tissues and Cardiovascular System modules
Both the male and the female reproductive systems consist of primary and secondary sex organs.
The primary sex organs are known as the gonads and consist of the testes in the male and the ovaries in the female.
The secondary sex organs are those that are necessary for the process of reproduction. In the male, these secondary sex organs consist of the ducts, the glands and the penis. In the female the uterine tubes (also called fallopian tubes), the uterus, and the vagina make-up the secondary organs.
Both the male and female sex organs are concerned with the process of uniting a spermatozoon (sperm), with the ovum in the process of fertilisation.
Both males and females have secondary sex characteristics which further distinguish males from females and are the characteristics which define puberty. These include: the growth of pubic and axillary hair in both sexes, facial hair, body hair and a change in the pitch of the voice in males, and in females the redistribution of body fat and the enlargement of the breasts.
The primary sex organs are known as the gonads and consist of the testes in the male and the ovaries in the female.
The secondary sex organs are those that are necessary for the process of reproduction. In the male, these secondary sex organs consist of the ducts, the glands and the penis. In the female the uterine tubes (also called fallopian tubes), the uterus, and the vagina make-up the secondary organs.
Both the male and female sex organs are concerned with the process of uniting a spermatozoon (sperm), with the ovum in the process of fertilisation.
Both males and females have secondary sex characteristics which further distinguish males from females and are the characteristics which define puberty. These include: the growth of pubic and axillary hair in both sexes, facial hair, body hair and a change in the pitch of the voice in males, and in females the redistribution of body fat and the enlargement of the breasts.
1 Male Anatomy and Physiology
|
|
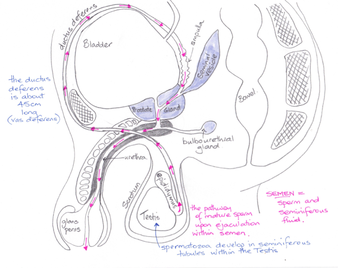
© Dr Patricia Logan 2017: male reproductive system. Note the pink arrows indicating the flow direction of semen
|
The male external genitalia consist of the scrotum and penis. The scrotum forms a pouch containing the testes and is important for maintaining the correct temperature for developing sperm. The scrotum is divided in to two by a septum; each houses one testicle. A bundle of fibrous tissue containing the ductus deferens, blood vessels, lymphatic vessels and nerves passes posteriorly from the scrotum, and is known as the spermatic cord.
The scrotum maintains the temperature of the testes at around 35° C, which is the ideal temperature for sperm production. Temperature maintenance is achieved by three mechanisms:
The testes are both endocrine and exocrine glands. The endocrine function is the production of sex hormones that are released to the bloodstream. The exocrine function describes the release of sperm through the ducts. The testes are surrounded by a white fibrous capsule from which connective tissue extends into the testes to form about 250 lobules. Each lobule contains one to three seminiferous tubules where sperm is produced. Lying between the seminiferous tubules are specialised cells known as Leydig cells (also called interstitial cells) that produce testosterone in response to the presence of luteinizing hormone (LH).
The seminiferous tubules are lined with several layers of germinal epithelium where sperm are found at different stages of development. Scattered amongst these germ cells are Sertoli cells (sustentacular cells) which protect the developing sperm by forming the blood-testes barrier and aid their development by providing nutrients and growth factors, removing waste, and releasing androgen-binding protein (ABP) and the hormone inhibin to regulate sperm production. The blood testes barrier prevents unwanted substances from entering the tubules. Germ cells are genetically different from other cells in the body and so the immune system may recognise these developing sperm as ‘non-self’ cells.
The seminiferous tubules converge on the rete testis, a network of tubes embedded in the capsule, where sperm continue to mature. At this stage in their development the sperm cannot swim and are dependent on fluid secreted by the Sertoli cells and cilia found on some of the rete cells to progress out of the testes to enter the epididymis. Most of the epididymis is composed a highly coiled structure through which sperm travel for about 20 days, slowly gaining the ability to swim, until they reach the tail of the epididymis. They are stored here until ejaculation. Stored sperm remain fertile for up to 60 days, at which time they are reabsorbed.
From the tail of the epididymis the sperm enter the ductus deferens (also called the vas deferens), a muscular tube about 45 cm in length. The ductus deferens travels up through the spermatic cord, passes between the bladder and ureters, before turning down behind the bladder and widening into the terminal ampulla where it then joins the seminal vesicle.
Three glands contribute fluid to the semen:
The penis consists of an internal root and an external portion, about 8 – 10 cm long. At the distal end of the penis is the glans, the head of the penis where the urethra exits the organ. There are three cylindrical layers of erectile tissue surrounding the penis, which become filled with blood during sexual arousal.
During ejaculation, approximately 2 – 5 ml semen is ejected from the penis and enters the vagina. Semen is a sticky fluid containing sperm, spermatic duct secretions, prostatic fluid, seminal vesicle fluid and a small amount of bulbourethral fluid. Sperm makes up about 10 % of the semen with 20 – 25 million sperm being present.
Once activated the sperm ‘swim’ through the female reproductive tract in search of an ovum to fertilise.
The scrotum maintains the temperature of the testes at around 35° C, which is the ideal temperature for sperm production. Temperature maintenance is achieved by three mechanisms:
- The cremaster muscle, which surrounds the spermatic cord, contracts or relaxes to move the testes either closer or further away from the body.
- The dartos muscle, surrounding the scrotum contracts when it is cold effectively wrinkling and tightening the scrotum reducing the surface area of the scrotum further reducing heat loss.
- A network of veins surround the testicular artery within the spermatic cord: as the arterial blood moves towards the testes, these veins which contain cooler blood, absorb the heat from the blood, so that when it reaches the testes it is about 2° C cooler than the core body temperature of 37° C.
The testes are both endocrine and exocrine glands. The endocrine function is the production of sex hormones that are released to the bloodstream. The exocrine function describes the release of sperm through the ducts. The testes are surrounded by a white fibrous capsule from which connective tissue extends into the testes to form about 250 lobules. Each lobule contains one to three seminiferous tubules where sperm is produced. Lying between the seminiferous tubules are specialised cells known as Leydig cells (also called interstitial cells) that produce testosterone in response to the presence of luteinizing hormone (LH).
The seminiferous tubules are lined with several layers of germinal epithelium where sperm are found at different stages of development. Scattered amongst these germ cells are Sertoli cells (sustentacular cells) which protect the developing sperm by forming the blood-testes barrier and aid their development by providing nutrients and growth factors, removing waste, and releasing androgen-binding protein (ABP) and the hormone inhibin to regulate sperm production. The blood testes barrier prevents unwanted substances from entering the tubules. Germ cells are genetically different from other cells in the body and so the immune system may recognise these developing sperm as ‘non-self’ cells.
The seminiferous tubules converge on the rete testis, a network of tubes embedded in the capsule, where sperm continue to mature. At this stage in their development the sperm cannot swim and are dependent on fluid secreted by the Sertoli cells and cilia found on some of the rete cells to progress out of the testes to enter the epididymis. Most of the epididymis is composed a highly coiled structure through which sperm travel for about 20 days, slowly gaining the ability to swim, until they reach the tail of the epididymis. They are stored here until ejaculation. Stored sperm remain fertile for up to 60 days, at which time they are reabsorbed.
From the tail of the epididymis the sperm enter the ductus deferens (also called the vas deferens), a muscular tube about 45 cm in length. The ductus deferens travels up through the spermatic cord, passes between the bladder and ureters, before turning down behind the bladder and widening into the terminal ampulla where it then joins the seminal vesicle.
Three glands contribute fluid to the semen:
- The seminal vesicles are a pair of glands lying posterior to the bladder. Each vesicle is associated with a ductus deferens. The ductus deferens and the duct from the seminal vesicle forms the ejaculatory duct which then passes through the prostate gland and into the urethra. The seminal vesicles add a yellowish fluid containing sugars to the semen. Sperm can use these sugars to make ATP for energy.
- The prostate gland lies inferior to the bladder and surrounds the urethra and ejaculatory ducts. This gland contributes a thin milky fluid to the semen. Prostate fluid elevates the pH to approximately 7.5 (alkaline) which is necessary to activate the sperm as they are ejaculated into the vagina.
- The bulbourethral glands are two pea-sized glands lying at the inner end of the penis, inferior to the prostate gland. These glands produce a clear, slippery fluid that serves to lubricate the penis during sexual arousal. More importantly, this fluid neutralises the acidity of any urine left in the urethra thus protecting the sperm.
The penis consists of an internal root and an external portion, about 8 – 10 cm long. At the distal end of the penis is the glans, the head of the penis where the urethra exits the organ. There are three cylindrical layers of erectile tissue surrounding the penis, which become filled with blood during sexual arousal.
During ejaculation, approximately 2 – 5 ml semen is ejected from the penis and enters the vagina. Semen is a sticky fluid containing sperm, spermatic duct secretions, prostatic fluid, seminal vesicle fluid and a small amount of bulbourethral fluid. Sperm makes up about 10 % of the semen with 20 – 25 million sperm being present.
Once activated the sperm ‘swim’ through the female reproductive tract in search of an ovum to fertilise.
2 Female anatomy and physiology
|
|
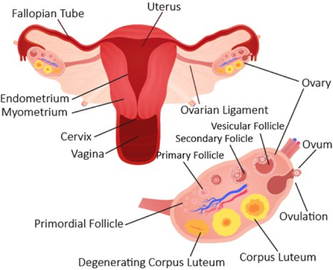
The female reproductive system highlighting the ovary (depositphotos.com)
|
The female is born with her lifetime supply of eggs which, by the time she reaches menopause (menstruation ceases), are no longer viable. The two ovaries, which produce the ova, are the primary sex organs of the female. The secondary sex organs include the fallopian tubes, the uterus and the vagina, and externally the clitoris, labia minora, and labia majora.
The ovary has a fibrous medulla where the veins and arteries that supply each ovary lie. The cortex contains the ovarian follicles which consist of one developing ova surrounded by smaller follicular cells. When an ovum reaches a certain stage of maturity the follicles burst through the ovary wall, the process termed ovulation, releasing the ovum towards the uterine (fallopian) tube. This occurs about once every 28 days, with usually only one ovum being released per ovarian cycle.
The uterine tubes are approximately 10 cm long and enable the ovum to reach the uterus. The wall of the tube contains smooth muscle with highly folded mucosa and ciliated epithelium. At the distal end of each fallopian tube are projections known as fimbriae, which sweep the ova from the surface of the ovary into the fallopian tube. The uterine tubes are the site of fertilisation.
This video from NewScientist shows the release of the ovum from the ovary and the fimbriae collecting it.
The ovary has a fibrous medulla where the veins and arteries that supply each ovary lie. The cortex contains the ovarian follicles which consist of one developing ova surrounded by smaller follicular cells. When an ovum reaches a certain stage of maturity the follicles burst through the ovary wall, the process termed ovulation, releasing the ovum towards the uterine (fallopian) tube. This occurs about once every 28 days, with usually only one ovum being released per ovarian cycle.
The uterine tubes are approximately 10 cm long and enable the ovum to reach the uterus. The wall of the tube contains smooth muscle with highly folded mucosa and ciliated epithelium. At the distal end of each fallopian tube are projections known as fimbriae, which sweep the ova from the surface of the ovary into the fallopian tube. The uterine tubes are the site of fertilisation.
This video from NewScientist shows the release of the ovum from the ovary and the fimbriae collecting it.
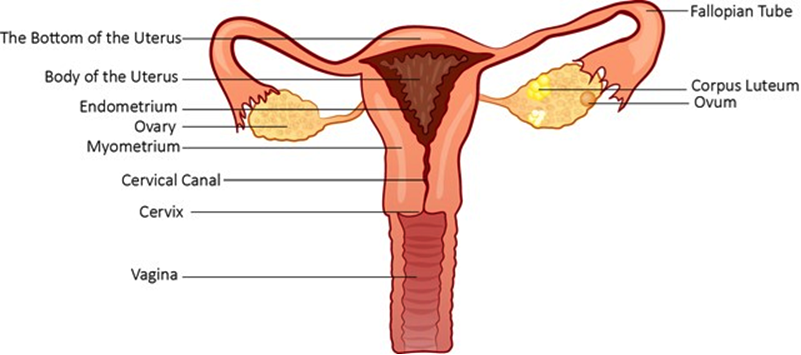
Structure of the female reproductive system (depositphotos.com)
The uterus is a muscular chamber that lies over the urinary bladder and opens into the vagina at the cervix. Its role is to protect the developing foetus, provide nutrition, and at the end of pregnancy to contract and expel the foetus. In a non-pregnant individual, the uterus is collapsed upon itself and occupies only a small amount of space.
The cervical canal forms the passageway between the uterus and the vagina; The superior opening being the internal os and the inferior opening the external os. Glands within the canal secrete mucous which helps to act as a barrier to microorganisms. At the time of ovulation (when an egg is released from the ovary), this mucous thins making an easier pathway for the entry and movement of sperm.
The vagina is an 8- 10 cm tube which serves as an entry for the penis and semen, the discharge of menstrual fluid and forms the birth canal. Its wall is thin but distensible and lubrication of the vagina occurs through the seepage of fluid through the vaginal wall and from cervical glands above it. In childhood, the vaginal epithelium is simple cuboidal cells, which with the release of oestrogen at puberty, transforms into stratified squamous epithelium (which is an example of metaplasia). Vaginal pH is low (acidic) at about 3.5, which along with immune cells in the mucosa, inhibits the growth of pathogens.
The external genitalia, known as the vulva, includes the mons pubis, labia majora, labia minora, clitoris, vaginal orifice and accessory glands.
The cervical canal forms the passageway between the uterus and the vagina; The superior opening being the internal os and the inferior opening the external os. Glands within the canal secrete mucous which helps to act as a barrier to microorganisms. At the time of ovulation (when an egg is released from the ovary), this mucous thins making an easier pathway for the entry and movement of sperm.
The vagina is an 8- 10 cm tube which serves as an entry for the penis and semen, the discharge of menstrual fluid and forms the birth canal. Its wall is thin but distensible and lubrication of the vagina occurs through the seepage of fluid through the vaginal wall and from cervical glands above it. In childhood, the vaginal epithelium is simple cuboidal cells, which with the release of oestrogen at puberty, transforms into stratified squamous epithelium (which is an example of metaplasia). Vaginal pH is low (acidic) at about 3.5, which along with immune cells in the mucosa, inhibits the growth of pathogens.
The external genitalia, known as the vulva, includes the mons pubis, labia majora, labia minora, clitoris, vaginal orifice and accessory glands.
The breast
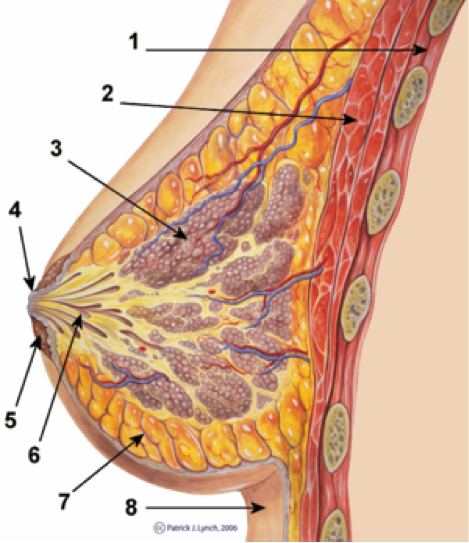
Original author: Patrick J. Lynch. Reworked by Morgoth666 to add numbered legend arrows. Creative Commons license. https://en.wikipedia.org/wiki/Breast#/media/File:Breast_anatomy_normal_scheme.png
|
The breast in cross-section:
|
The breast lies over the pectoralis major muscle and is a tissue that enlarges at puberty. It consists of adipose and collagenous tissue in the non-pregnant female, with a series of ducts which converge on the nipple. The mammary glands develop during pregnancy and are active throughout the lactation period, atrophying when breastfeeding finishes. The breast becomes more lobular during pregnancy, with about 15 -20 lobes surrounding each nipple. These lobes are drained by the lactiferous ducts which widen into lactiferous sinuses entering the nipple. At the distal end each duct branches into numerous sacs known as acini, surrounded by myoepithelial cells which are involved in the release of milk.
3 Cell Division: Mitosi and Meiosis
Mitosis and meiosis are the mechanisms the body uses to create new cells. In mitosis the daughter cells are identical to the parent cell whereas in meiosis, the new cells are non-identical and contain only half the number of chromosomes.
Nearly all cells contain deoxyribonucleic acid (DNA) within their nucleus. DNA is made up of coded sequences known as genes. Genes are the instructions for making proteins, and are the units of heredity. When the instruction carried by the DNA has an error in it, a mutated gene, then the result can be conditions such as muscular dystrophy or cystic fibrosis. Within the nucleus, DNA is found as thread-like structures known as chromatin when the cell is not undergoing division.
Mitosis is the mechanism whereby individual cells replicate their DNA and other cellular structures in order to split into two identical daughter cells. Cells undergo mitosis in order to replace themselves such as when the epithelial lining of the gut is replaced roughly every three days or to effect tissue repair after trauma.
Meiosis is the process that enables the chromosome count within germ cells, those that form sperm and ovum (egg), to be reduced by half so that when egg and sperm meet the chromosome count remains the same as the parent’s in the offspring.
In human cells, there are 46 chromosomes found in pairs (23 pairs), 22 pairs are autosomes and there is one pair of sex chromosomes, XX for a female and XY for a male. Each pair of chromosomes carries the same genes, although there may be slight variations in the gene. For example, the gene for eye colour may code for blue on one chromosome and green on the other. These pairs of genes are known as alleles.
Nearly all cells contain deoxyribonucleic acid (DNA) within their nucleus. DNA is made up of coded sequences known as genes. Genes are the instructions for making proteins, and are the units of heredity. When the instruction carried by the DNA has an error in it, a mutated gene, then the result can be conditions such as muscular dystrophy or cystic fibrosis. Within the nucleus, DNA is found as thread-like structures known as chromatin when the cell is not undergoing division.
Mitosis is the mechanism whereby individual cells replicate their DNA and other cellular structures in order to split into two identical daughter cells. Cells undergo mitosis in order to replace themselves such as when the epithelial lining of the gut is replaced roughly every three days or to effect tissue repair after trauma.
Meiosis is the process that enables the chromosome count within germ cells, those that form sperm and ovum (egg), to be reduced by half so that when egg and sperm meet the chromosome count remains the same as the parent’s in the offspring.
In human cells, there are 46 chromosomes found in pairs (23 pairs), 22 pairs are autosomes and there is one pair of sex chromosomes, XX for a female and XY for a male. Each pair of chromosomes carries the same genes, although there may be slight variations in the gene. For example, the gene for eye colour may code for blue on one chromosome and green on the other. These pairs of genes are known as alleles.
Mitosis
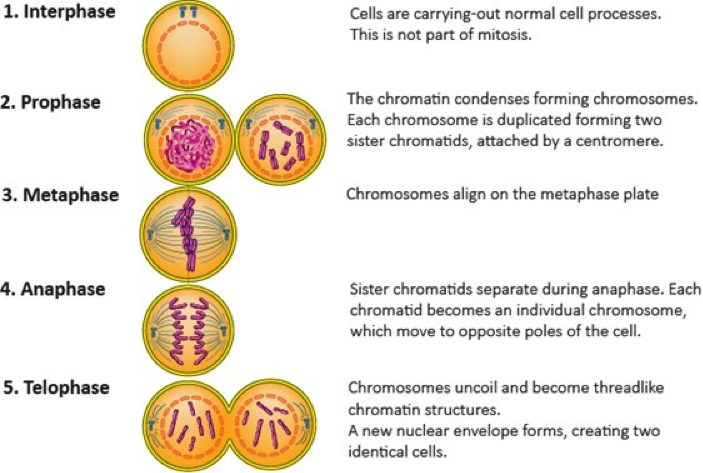
The stages of mitosis (depositphotos.com)
During mitosis and meiosis, the chromatin condenses and forms segments of different lengths known as chromosomes.
The phases of mitosis are labelled:
Prophase - chromatin condenses into chromosomes, duplicated along their length;
Metaphase - chromosomes line up on the mitotic spindle;
Anaphase - the sister chromatids are pulled to either end of the cell;
Telophase - chromosomes spread back out into chromatin threads and a new nuclear envelope encases them at either end of the dividing cell; and finally ...
Cytokinesis - where the cell divides in two.
The phases of mitosis are labelled:
Prophase - chromatin condenses into chromosomes, duplicated along their length;
Metaphase - chromosomes line up on the mitotic spindle;
Anaphase - the sister chromatids are pulled to either end of the cell;
Telophase - chromosomes spread back out into chromatin threads and a new nuclear envelope encases them at either end of the dividing cell; and finally ...
Cytokinesis - where the cell divides in two.
Meiosis
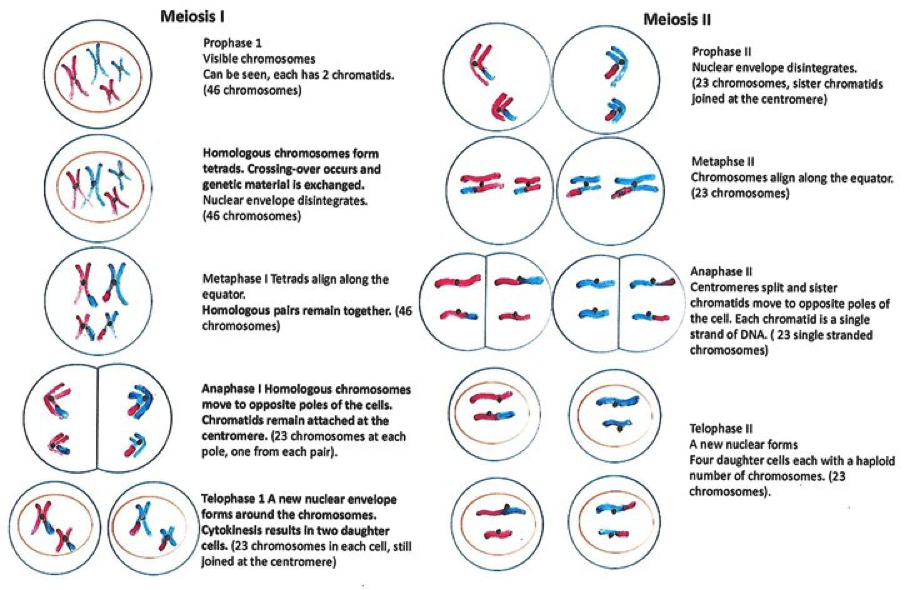
The stages of meiosis. (adapted from Saladin. 2015)
Meiosis occurs in germ cells and is comprised of two overarching phases: meiosis 1 – which resembles mitosis, and meiosis 2. The germ cells are those that become the ovum (pl. ova) or the spermatozoa (sing. spermatozoon). The end-product of meiosis is four non-identical cells which contain 23 single chromosomes instead of 46 (23 pairs). Once an ovum combines with a sperm at fertilisation, the chromosome count in the combined cell is returned to 46.
Males constantly produce new sperm cells from the time of puberty. Sperm production is termed spermatogenesis.
In females, the development of ova is begun before birth and then suspended until puberty when meiosis I is completed. One of the two cells produced will disintegrate (termed a polar body) while the other, now called a secondary oocyte, continues to metaphase 2. Ova (egg) production is termed oogenesis. This cell will not complete meiosis unless it is fertilised. At fertilisation, the continuing cell division results in a second polar body being produced which again disintegrates, while the larger remaining ovum combines its DNA with that of the sperm cell.
Males constantly produce new sperm cells from the time of puberty. Sperm production is termed spermatogenesis.
In females, the development of ova is begun before birth and then suspended until puberty when meiosis I is completed. One of the two cells produced will disintegrate (termed a polar body) while the other, now called a secondary oocyte, continues to metaphase 2. Ova (egg) production is termed oogenesis. This cell will not complete meiosis unless it is fertilised. At fertilisation, the continuing cell division results in a second polar body being produced which again disintegrates, while the larger remaining ovum combines its DNA with that of the sperm cell.
4 Spermatogenesis and Oogenesis
|
|
|
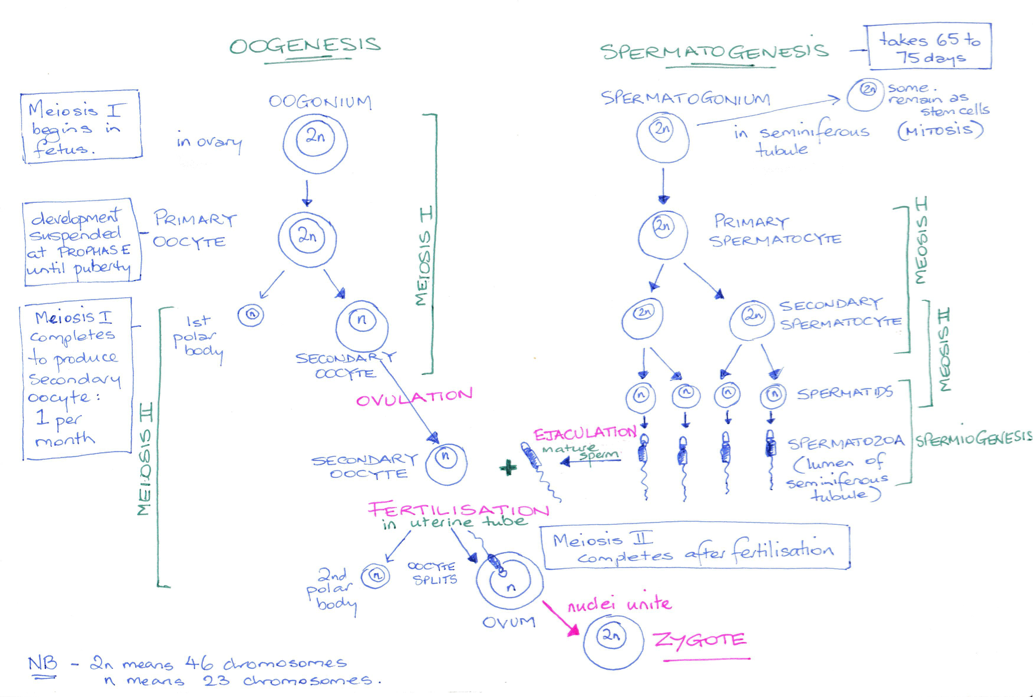
© Dr Patricia Logan 2017
Spermatogenesis and oogenesis are terms used to describe the development of the sperm and the egg respectively. While both processes result in a gamete containing a haploid (only one copy of each), number of chromosomes, the events that occur are significantly different in that the male produces four haploid daughter cells (sperm), and the female only produces one viable daughter cell (the ovum).
Spermatogenesis takes place in the seminiferous tubules with the developing gametes moving from the basement membrane towards the lumen of the tubules as meiosis proceeds. By the time the gametes reach the lumen immature sperm are present.
The first stem cells which develop into sperm cells are known as primordial germ cells. These cells form in the developing embryo and colonise the gonadal ridges by about six weeks post conception. Now known as spermatogonia, they remain dormant until puberty and spend their time occupying a space close to the basement membrane. At puberty, when testosterone levels rise, these spermatogonia become activated and spermatogenesis commences.
Spermatogenesis takes place in the seminiferous tubules with the developing gametes moving from the basement membrane towards the lumen of the tubules as meiosis proceeds. By the time the gametes reach the lumen immature sperm are present.
The first stem cells which develop into sperm cells are known as primordial germ cells. These cells form in the developing embryo and colonise the gonadal ridges by about six weeks post conception. Now known as spermatogonia, they remain dormant until puberty and spend their time occupying a space close to the basement membrane. At puberty, when testosterone levels rise, these spermatogonia become activated and spermatogenesis commences.
Spermatogenesis
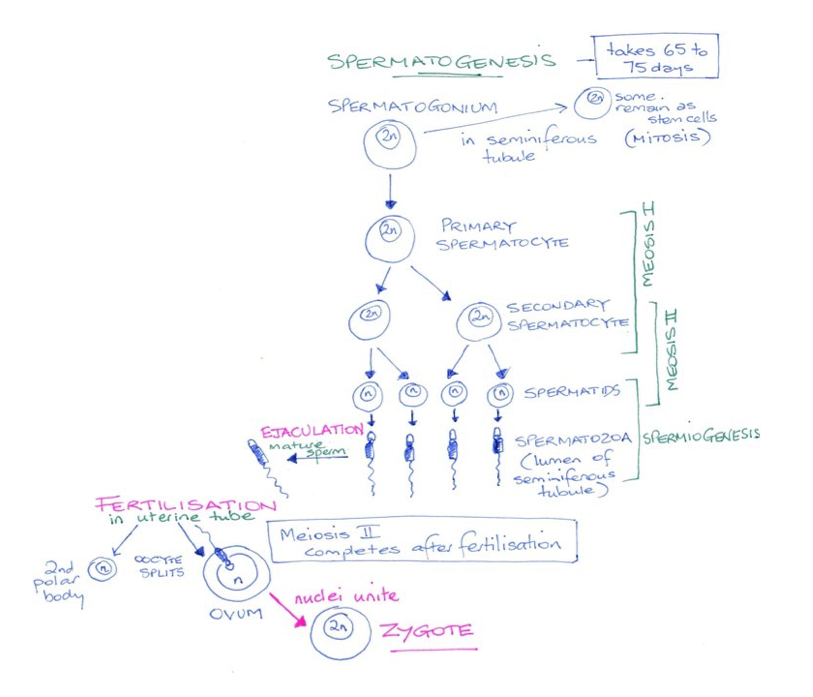
- Spermatogonia divide by mitosis, producing two diploid daughter cells. One of these cells remains close to the basement membrane where it provides a lifetime supply of stem cells for the production of sperm.
- The other daughter cell moves away from the basement membrane towards the lumen. These cells grow and become primary spermatocytes.
- Tight junctions between the sertoli cells break-down to allow these spermatocytes through to the lumen and then they reform. The sertoli cells protect the spermatocytes from immune recognition as they undergo meiosis and become genetically different from the rest of the body cells.
- The primary spermatocytes undergo meiosis I resulting in two haploid genetically unique secondary spermatocytes.
- Each secondary spermatocyte now undergoes meiosis II; the sister chromatids separate, a new cell membrane is formed resulting in four spermatids. For each spermatogonia four genetically unique spermatids result.
- The spermatid now undergoes a process of spermiogenesis - it grows a flagella (tail) and it releases most of its cytoplasm, making it lighter. It enters the lumen of the seminiferous tubule, and continues to mature as it makes its way towards the tail of the epididymis.
Oogenesis
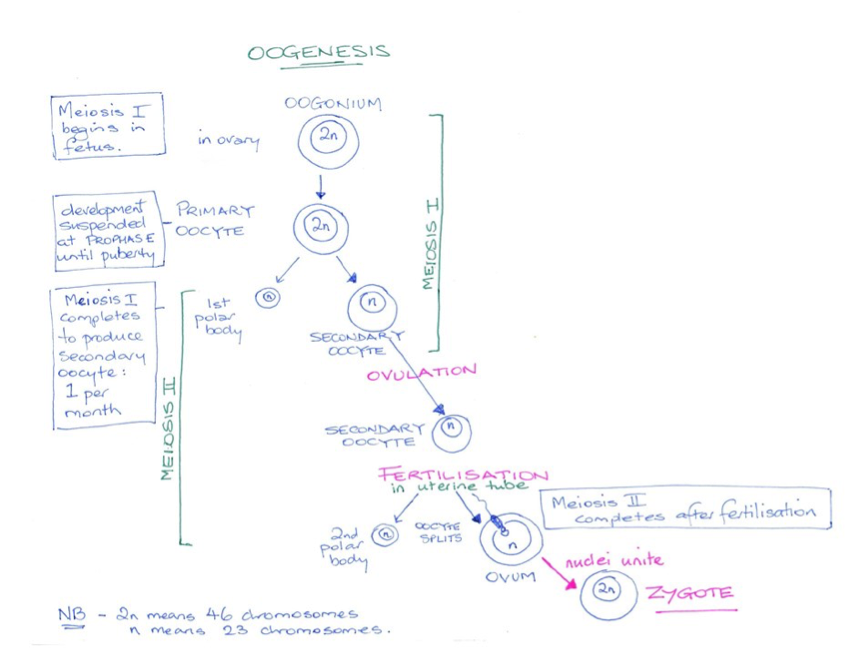
© Dr Patricia Logan 2017
- The first germ cells migrate to the gonadal ridges in the first six weeks of embryonic development and differentiate into oogonia.
- Oogonia increase in number during the first four to five months of fetal development resulting in approximately six million cells. Development then stops until just prior to birth.
- During the latter stages of the mother’s pregnancy, the fetus’s oogonia transform into primary oocytes and go through early meiosis I.
- By birth no oogonia remain and most primary oocytes degenerate with only about two million remaining at birth.
- Throughout childhood, puberty and through to menopause many more primary oocytes are lost. At puberty about 200,000 primary oocytes remain. This is a woman’s reproductive supply to last through her fertile years.
From puberty until menopause several arrested follicles recommence development. Over a period of 290 days a follicle will develop to a fully mature ovum and be released from the ovary. Meiosis 1 is completed at the time of ovulation resulting in two daughter cells. One of these is released as a large secondary oocyte while the other much smaller cell lacks cytoplasm and is known as the first polar body. This polar body degenerates. The secondary oocyte continues through meiosis II until metaphase at which time the process pauses and will not proceed further unless the egg is fertilised by a sperm. If fertilised the ovum will complete meiosis II, producing a second polar body that also disintegrates. If not fertilised, the ovum fails to develop further and is lost from the body during menstruation.
Follicle development
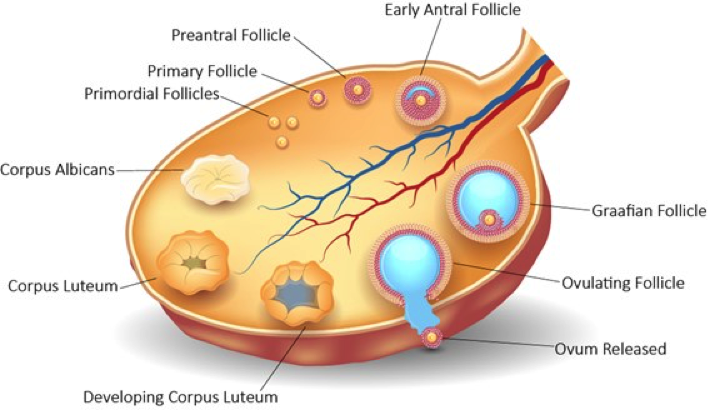
Stages of follicular development in the ovary (depositphotos.com)
Several stages of follicle development occur leading to ovulation of the ovum. These are briefly described below.
- A primordial follicle containing the primary oocyte in the early stages of meiosis 1 becomes activated. Usually about two dozen primary oocytes are activated at any one time, but ordinarily only one will make it through the 290-day development cycle and be ovulated.
- At about 140 days: the primordial follicle will have become a primary follicle containing a larger secondary oocyte. Growth factors are involved in this process and the follicular cells surrounding the follicle become granulosa cells that develop follicle stimulating hormone (FSH) receptors. These require stimulation by FSH to progress the primary follicle to become secondary follicles.
- At about 170 days: Secondary follicles appear and are larger than primary follicles having several layers of granulosa cells surrounding the follicle. The oocyte is surrounded by a layer of protein gel called the zona pellucida. The outer layer of the follicle has now developed a thick hardened layer known as the theca folliculi, which contains a supply of blood vessels delivering nutrition, cholesterol and hormones to the secondary follicle.
- At about 60 days: before ovulation the granulosa cells start to secrete follicular fluid to form the antrum. This is now a tertiary (antral or vesicular) follicle. The innermost layer of these granulosa cells, surrounding the zona pellucida and the oocyte, is known as the corona radiata. The oocyte and the corona radiata grow microvilli, and gap junctions form between the cells allowing the passage of nutrients and waste. The egg is now well protected and cannot be penetrated unless the corona radiata is breached.
- At about 20 days prior to ovulation: one follicle becomes dominant and will be ovulated. Although not fully understood, this follicle manages to sequester most of the FSH and continues to grow while the growth rate in the other tertiary follicles slows. By 5 days pre-ovulation this dominant follicle is about 20 mm in diameter and is known as a Graafian follicle which can be seen on ultrasound examination. At the time of ovulation, it has grown to 30 mm in diameter and the oocyte is floating freely in the antrum.
5 The ovarian and uterine cycle
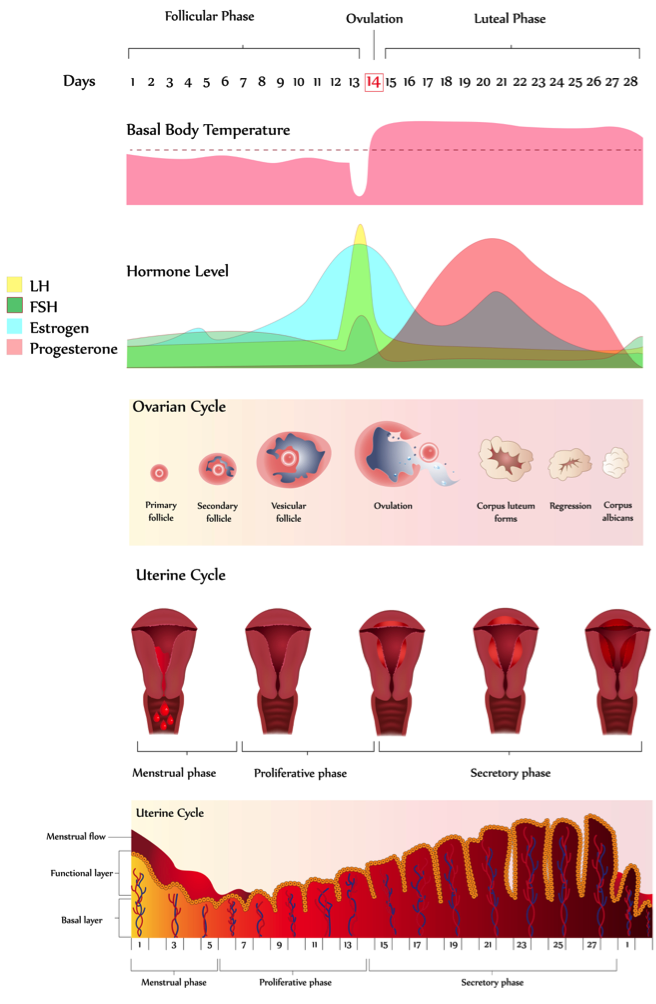
The ovarian cycle (depositphotos.com)
The ovarian cycle lasts approximately 28 days, although it varies from individual to individual, and can vary month to month. The cycle is regulated by the hypothalamus-pituitary-ovarian axis. Hormones from the hypothalamus regulate the pituitary gland, which in turn regulates the ovaries, and the ovaries in turn regulate the uterus.
Days 1 -5 (approximately) are the menstrual cycle where blood and endometrial tissue is discharged. At the same time, a two-week follicular cycle begins when FSH stimulates the growth of follicles and in particular the growth of the dominant follicle. Granulosa cells secrete oestrogen, which upregulates FSH receptors, oestrogen receptors and luteinising hormone (LH) receptors on the granulosa cells of the dominant follicle. A rise in all three hormones will be observed. Oestrogen also inhibits the hypothalamus from releasing gonadotropin releasing hormone (GnRH), influencing the pituitary gland to release less FSH and more LH.
Cell mitosis replaces the lost uterine tissue; a process that takes approximately 20 days. During this phase tertiary follicles grow until around day 14 when ovulation occurs and the dominant follicle releases its ovum. As ovulation approaches oestrogen stimulates a surge in LH and a spike in FSH secretion by the pituitary gland. The primary oocyte completes meiosis 1 resulting in the first polar body and a haploid secondary oocyte.
Following ovulation, the cycle enters the luteal phase. The remainder of the dominant follicle becomes the corpus luteum, regulated by LH. This hormone stimulates the corpus luteum to secrete oestrogen and progesterone. Progesterone plays a crucial role in preparing the uterus for pregnancy by stimulating the endometrium to thicken and then maintaining it if fertilisation occurs until around 12 weeks of pregnancy. High levels of oestrogen, progesterone, and inhibin from the corpus luteum have an inhibitory effect on the pituitary leading to declining levels of LH and FSH. This is the method by which some contraceptive pills control the cycle. If no fertilisation occurs, then the corpus luteum shrinks and by day 26 of the cycle has become scar tissue, known as the corpus albicans. The endometrium breaks down as the hormone levels fall and the accumulation of tissue and blood results in menstruation. As the levels of progesterone and oestrogen decline then the pituitary is no longer inhibited. It starts to secrete FSH stimulating the ovarian follicles and the cycle begins again.
Days 1 -5 (approximately) are the menstrual cycle where blood and endometrial tissue is discharged. At the same time, a two-week follicular cycle begins when FSH stimulates the growth of follicles and in particular the growth of the dominant follicle. Granulosa cells secrete oestrogen, which upregulates FSH receptors, oestrogen receptors and luteinising hormone (LH) receptors on the granulosa cells of the dominant follicle. A rise in all three hormones will be observed. Oestrogen also inhibits the hypothalamus from releasing gonadotropin releasing hormone (GnRH), influencing the pituitary gland to release less FSH and more LH.
Cell mitosis replaces the lost uterine tissue; a process that takes approximately 20 days. During this phase tertiary follicles grow until around day 14 when ovulation occurs and the dominant follicle releases its ovum. As ovulation approaches oestrogen stimulates a surge in LH and a spike in FSH secretion by the pituitary gland. The primary oocyte completes meiosis 1 resulting in the first polar body and a haploid secondary oocyte.
Following ovulation, the cycle enters the luteal phase. The remainder of the dominant follicle becomes the corpus luteum, regulated by LH. This hormone stimulates the corpus luteum to secrete oestrogen and progesterone. Progesterone plays a crucial role in preparing the uterus for pregnancy by stimulating the endometrium to thicken and then maintaining it if fertilisation occurs until around 12 weeks of pregnancy. High levels of oestrogen, progesterone, and inhibin from the corpus luteum have an inhibitory effect on the pituitary leading to declining levels of LH and FSH. This is the method by which some contraceptive pills control the cycle. If no fertilisation occurs, then the corpus luteum shrinks and by day 26 of the cycle has become scar tissue, known as the corpus albicans. The endometrium breaks down as the hormone levels fall and the accumulation of tissue and blood results in menstruation. As the levels of progesterone and oestrogen decline then the pituitary is no longer inhibited. It starts to secrete FSH stimulating the ovarian follicles and the cycle begins again.
6 Pregnancy, development and birth
Fertilisation
The egg can survive for up to 24 h post-ovulation but takes up to 72 h to reach the uterus. It therefore needs to meet the sperm while travelling through the uterine tube if it is to be fertilised. Sperm travel through the female reproductive tract and can reach the distal end of the fallopian tube within 0.5 h post-ejaculation, but are incapable of fertilising the egg for another 10 h, during which time they undergo a transformation known as capacitation.
On encountering an ovum the sperm undergoes an acrosomal reaction whereby the acrosome releases enzymes which allow penetration of the egg. It takes several sperm to breach the granulosa cells and zona pellucida. Once a sperm makes contact with the egg the other sperm are locked out. Only the sperm head and midpiece enter the egg, which destroys the sperm’s mitochondria, (mitochondrial DNA found in a person’s cells is maternal DNA only). Once the egg is penetrated meiosis II is completed and the diploid number of chromosomes is reconstituted with half the chromosomes coming from the male and half from the female. This diploid cell is now known as the zygote, and will continue to divide by mitosis.
On encountering an ovum the sperm undergoes an acrosomal reaction whereby the acrosome releases enzymes which allow penetration of the egg. It takes several sperm to breach the granulosa cells and zona pellucida. Once a sperm makes contact with the egg the other sperm are locked out. Only the sperm head and midpiece enter the egg, which destroys the sperm’s mitochondria, (mitochondrial DNA found in a person’s cells is maternal DNA only). Once the egg is penetrated meiosis II is completed and the diploid number of chromosomes is reconstituted with half the chromosomes coming from the male and half from the female. This diploid cell is now known as the zygote, and will continue to divide by mitosis.
Foetal development
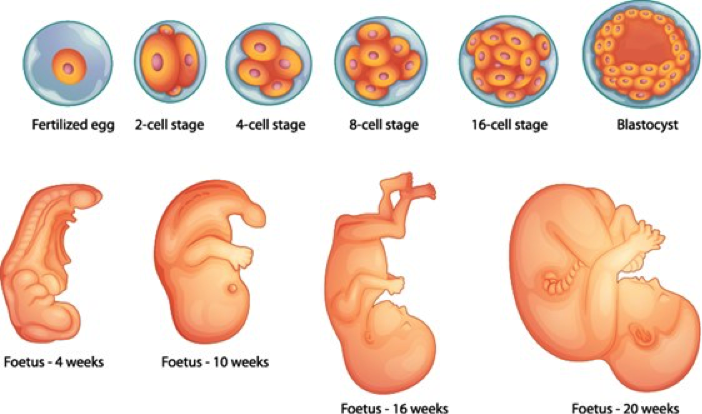
Foetal development during the first 20 weeks post conception. (Depositphotos.com)
The products of conception – the embryo or foetus plus the placental membranes - are termed a conceptus. The conceptus is a solid ball of cells named a morula for the first 4 to 5 days and then develops a fluid filled cavity at which time it is termed a blastocyst. A thickened area of cells in the blastocyst form the inner cell mass which will become the embryo proper. The blastocyst embeds in to the uterine wall around 7 days after fertilisation.
From weeks 2 to 8 it is known as an embryo and from the 9th week until birth it is known as a foetus.
The umbilical cord, made up of two veins and one artery, connect the foetus to the placenta, which is attached to the uterine wall. The placenta acts as an endocrine gland, taking over the role from the corpus luteum, releasing hormones to help maintain the pregnancy as well as enabling gas exchange and waste removal. There is no blood exchanged between the mother and the foetus.
Pregnancy is divided into trimesters with the first trimester lasting from fertilisation until 12 weeks. This stage of pregnancy presents the greatest risk to the developing foetus, with up to 50% of pregnancies failing during this stage. The second trimester covers weeks 13 – 24, and is the time when most organ development is completed. From 25 weeks until birth the foetus continues to grow and most organs are matured to support life outside the womb.
Several hormones interact during pregnancy of which oestrogen, progesterone, human chorionic gonadatrophin (HCG). These important hormones are secreted initially by the corpus luteum (until it degenerates) or the blastocyst. From week 7 - 17 the placenta becomes the major source of hormones.
HCG levels rise until about week 12 post-conception then fall to low levels for the remainder of the pregnancy. HCG from the blastocyst stimulates the growth of the corpus luteum which increases its secretion of progesterone and oestrogen. It is the presence of raised HCG levels that form the basis of the urinary and blood pregnancy tests.
Oestrogen levels rise throughout pregnancy and stimulate the growth of the foetus and of the mother. The woman’s uterus enlarges, mammary ducts grow, the pubic symphysis becomes more elastic, and the sacroiliac joints loosen in preparation for childbirth.
Large amounts of progesterone are secreted in the early stages of pregnancy, inhibiting the production of FSH and LH. Progesterone stops the uterus from contracting, stops menstruation, and stimulates the growth of decidual cells, (the blastocysts nutrient supply), of the endometrium. Following the growth of mammary ducts stimulated by oestrogen, progesterone stimulates the growth of secretory acini, in preparedness for breastfeeding.
From weeks 2 to 8 it is known as an embryo and from the 9th week until birth it is known as a foetus.
The umbilical cord, made up of two veins and one artery, connect the foetus to the placenta, which is attached to the uterine wall. The placenta acts as an endocrine gland, taking over the role from the corpus luteum, releasing hormones to help maintain the pregnancy as well as enabling gas exchange and waste removal. There is no blood exchanged between the mother and the foetus.
Pregnancy is divided into trimesters with the first trimester lasting from fertilisation until 12 weeks. This stage of pregnancy presents the greatest risk to the developing foetus, with up to 50% of pregnancies failing during this stage. The second trimester covers weeks 13 – 24, and is the time when most organ development is completed. From 25 weeks until birth the foetus continues to grow and most organs are matured to support life outside the womb.
Several hormones interact during pregnancy of which oestrogen, progesterone, human chorionic gonadatrophin (HCG). These important hormones are secreted initially by the corpus luteum (until it degenerates) or the blastocyst. From week 7 - 17 the placenta becomes the major source of hormones.
HCG levels rise until about week 12 post-conception then fall to low levels for the remainder of the pregnancy. HCG from the blastocyst stimulates the growth of the corpus luteum which increases its secretion of progesterone and oestrogen. It is the presence of raised HCG levels that form the basis of the urinary and blood pregnancy tests.
Oestrogen levels rise throughout pregnancy and stimulate the growth of the foetus and of the mother. The woman’s uterus enlarges, mammary ducts grow, the pubic symphysis becomes more elastic, and the sacroiliac joints loosen in preparation for childbirth.
Large amounts of progesterone are secreted in the early stages of pregnancy, inhibiting the production of FSH and LH. Progesterone stops the uterus from contracting, stops menstruation, and stimulates the growth of decidual cells, (the blastocysts nutrient supply), of the endometrium. Following the growth of mammary ducts stimulated by oestrogen, progesterone stimulates the growth of secretory acini, in preparedness for breastfeeding.
Maternal changes during pregnancy
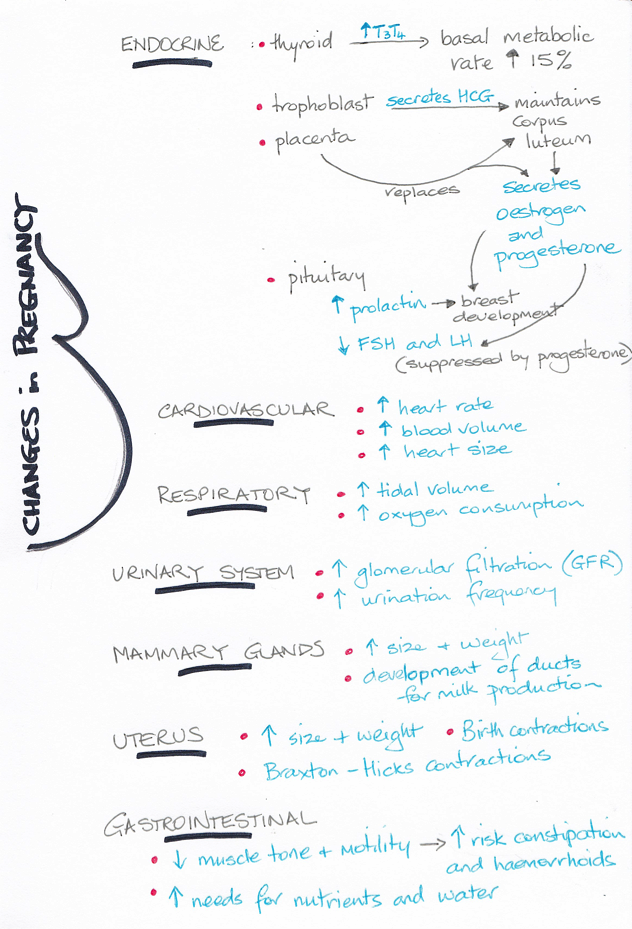
© Dr Patricia Logan 2017
The female anatomy and physiology has to make several adjustments in order to support the developing foetus and prepare for childbirth and supporting the baby following birth. Some of these changes have been mentioned above. Below is a list of other changes observed:
During the first six weeks after birth, the mother’s anatomy and physiology returns to its pre-pregnancy (pregravid) state. The uterus shrinks through autolysis of uterine cells. This is encouraged by breast-feeding, which assists in keeping oestrogen levels low and maintains the secretion of oxytocin.
Milk synthesis is promoted by the anterior pituitary hormone prolactin and starts initially with the release of colostrum - a substance similar to breast milk but containing about 30 % less fat. This substance contains immunoglobulins, in particular IgA, which protects the infant against gastroenteritis. Prolactin secretion is stimulated by breastfeeding. Between feeds this hormone returns to baseline levels.
- Morning sickness, although the reasons for it are not fully understood;
- The mother’s blood volume increases by about 30 % during pregnancy and cardiac output increases by up to 40 % by the end of the second trimester;
- While the respiratory rate remains stable throughout the pregnancy, tidal volume and minute ventilation increase by about 40 %;
- From about 4.5 months, the woman’s basal metabolic rate increases by about 15 %;
- Constipation may result from decreased intestinal motility;
- Heartburn is common during pregnancy due to the pressure of the growing uterus on the stomach;
- While aldosterone and steroid hormones released during pregnancy promote water retention and the glomerular filtration rate (GFR) increases by up to 50 %, diuresis is increased only slightly.
- The stretching of the dermis as the skin (integument) layer grows can result in tears to the connective tissue leading to striae (stretch marks):
- The skin of the abdomen grows to accommodate the expanding uterus;
- The skin covering the breasts grows to accommodate their enlargement.
During the first six weeks after birth, the mother’s anatomy and physiology returns to its pre-pregnancy (pregravid) state. The uterus shrinks through autolysis of uterine cells. This is encouraged by breast-feeding, which assists in keeping oestrogen levels low and maintains the secretion of oxytocin.
Milk synthesis is promoted by the anterior pituitary hormone prolactin and starts initially with the release of colostrum - a substance similar to breast milk but containing about 30 % less fat. This substance contains immunoglobulins, in particular IgA, which protects the infant against gastroenteritis. Prolactin secretion is stimulated by breastfeeding. Between feeds this hormone returns to baseline levels.
7 Inheritance
Heredity is the study of the inheritance of genetic traits. Genetic material constitutes genes, and genes are lengths of DNA that code in the most part for proteins. Some traits are determined by a single gene and are known as Mendelian traits. Some traits are determined by more than one gene and these are known as polygenic traits. For ease of understanding we will only focus on Mendelian inheritance here.
Humans have 46 chromosomes, with two copies of chromosomes 1 – 22 and then either a pair of X chromosomes or an X and a Y. Each pair are known as homologous chromosomes and they contain the same genes at the same position on each chromosome. These pairs of genes are known as alleles: one is inherited from the mother and one from the father. The genes that a person carries are known as the person’s genotype. Note that in the case of the male, the sex chromosomes are X and Y and so these are a non-homologous pair and do not contain the same genes. They do not crossover during meiosis I.
Inheritance is termed either autosomal inheritance or sex-linked inheritance. Autosomal inheritance concerns the genes on chromosomes 1 to 22 while sex-linked inheritance concerns the genes on the X and Y chromosome.
While each allele of a gene codes for the same trait, there may be slight differences in the coding sequence of the gene. This means that one may be expressed in the person and the other may not be seen. The expressed allele (that one that is visible or evident on testing) is known as the dominant gene, and the other is known as the recessive gene. The dominant one in the pair will be expressed. A person with two dominant genes will express the trait - they have a homozygous (two the same) pair of genes for the trait. A person who is heterozygous for the trait, that is they have one dominant and one recessive gene, will only express the dominant gene. For the recessive gene to be visible a person needs to possess two of the recessive genes – ie. Be homozygous recessive for the trait. The trait that is expressed is termed the phenotype. Examples of phenotypes are brown hair or blue eyes, or blood type A.
To add to the complexity of genetic variation, some traits are neither dominant nor recessive. This is known as co-dominance. The alleles are heterozygous but both are expressed giving an intermediatory phenotype. This is the case with the ABO blood group, which we will look at shortly.
Humans have 46 chromosomes, with two copies of chromosomes 1 – 22 and then either a pair of X chromosomes or an X and a Y. Each pair are known as homologous chromosomes and they contain the same genes at the same position on each chromosome. These pairs of genes are known as alleles: one is inherited from the mother and one from the father. The genes that a person carries are known as the person’s genotype. Note that in the case of the male, the sex chromosomes are X and Y and so these are a non-homologous pair and do not contain the same genes. They do not crossover during meiosis I.
Inheritance is termed either autosomal inheritance or sex-linked inheritance. Autosomal inheritance concerns the genes on chromosomes 1 to 22 while sex-linked inheritance concerns the genes on the X and Y chromosome.
While each allele of a gene codes for the same trait, there may be slight differences in the coding sequence of the gene. This means that one may be expressed in the person and the other may not be seen. The expressed allele (that one that is visible or evident on testing) is known as the dominant gene, and the other is known as the recessive gene. The dominant one in the pair will be expressed. A person with two dominant genes will express the trait - they have a homozygous (two the same) pair of genes for the trait. A person who is heterozygous for the trait, that is they have one dominant and one recessive gene, will only express the dominant gene. For the recessive gene to be visible a person needs to possess two of the recessive genes – ie. Be homozygous recessive for the trait. The trait that is expressed is termed the phenotype. Examples of phenotypes are brown hair or blue eyes, or blood type A.
To add to the complexity of genetic variation, some traits are neither dominant nor recessive. This is known as co-dominance. The alleles are heterozygous but both are expressed giving an intermediatory phenotype. This is the case with the ABO blood group, which we will look at shortly.
Simple Mendelian Inheritance
Here we will look at single gene traits as examples for inheritance patterns and the probability of inheriting a particular genetic trait. Please note that in humans the inheritance of different traits is often more complex than these examples and involve more than one gene. We can determine the genotype and phenotype of offspring if we know the genotype of the parents. The use of punnet squares assists in our developing an understanding of inheritance. In a punnet square the genotype of one parent is entered along the horizontal axis of the punnet square and the genotype of the other parent is entered along the vertical axis. Each allele is entered in a separate column or row, leading to a grid pattern of combinations for the specific trait.
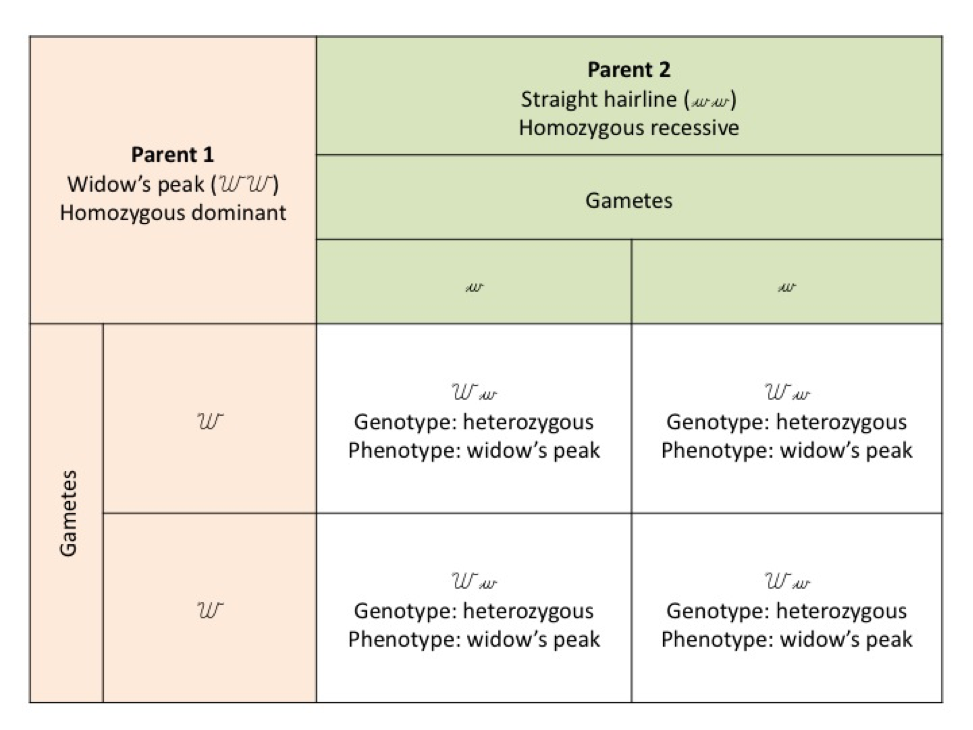
Punnet square A for autosomal inheritance where one parent is homozygous dominant and one parent is homozygous recessive for widow’s peak.
To interpret punnet square A, firstly look at the parental genotypes. One parent has a widow’s peak (a point in the centre of the hairline) with two dominant alleles (we will use WW) and the other parent has a straight hairline, with two recessive alleles (ww). When the gametes come together at fertilisation the offspring can only receive a widow’s peak dominant allele from parent 1 and a straight hairline recessive allele from parent 2.
All offspring will be heterozygous for the dominant gene and will have a widow’s peak (Ww).
All offspring will be heterozygous for the dominant gene and will have a widow’s peak (Ww).
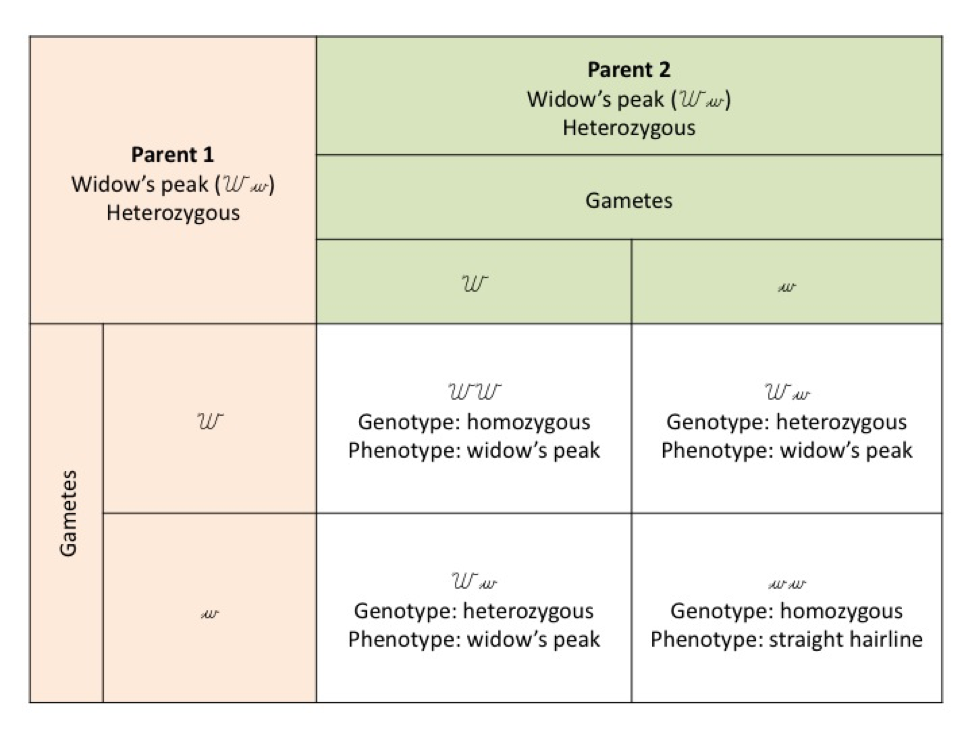
Punnet square B showing autosomal inheritance of the widow’s peak when both parents are heterozygous for the gene
In punnet square B above both parents are heterozygous for widow’s peak (Ww). At fertilisation the offspring can receive both dominant genes (one from each parent) WW, a dominant gene from one parent and a recessive from the other (Ww), or both recessive genes (one from each parent), (ww). Where both parents are heterozygous for the recessive allele – the chances for a straight hairline in the offspring becomes 1 in 4.
All three genotypes are present, but notice that there are two phenotypes, either widow’s peak present or a straight hairline.
From these outcomes, you should be able to see that if both parents have a straight hairline (ww) then only the recessive allele can be passed on to the offspring and all offspring will have a straight hairline.
All three genotypes are present, but notice that there are two phenotypes, either widow’s peak present or a straight hairline.
From these outcomes, you should be able to see that if both parents have a straight hairline (ww) then only the recessive allele can be passed on to the offspring and all offspring will have a straight hairline.
Co-dominance
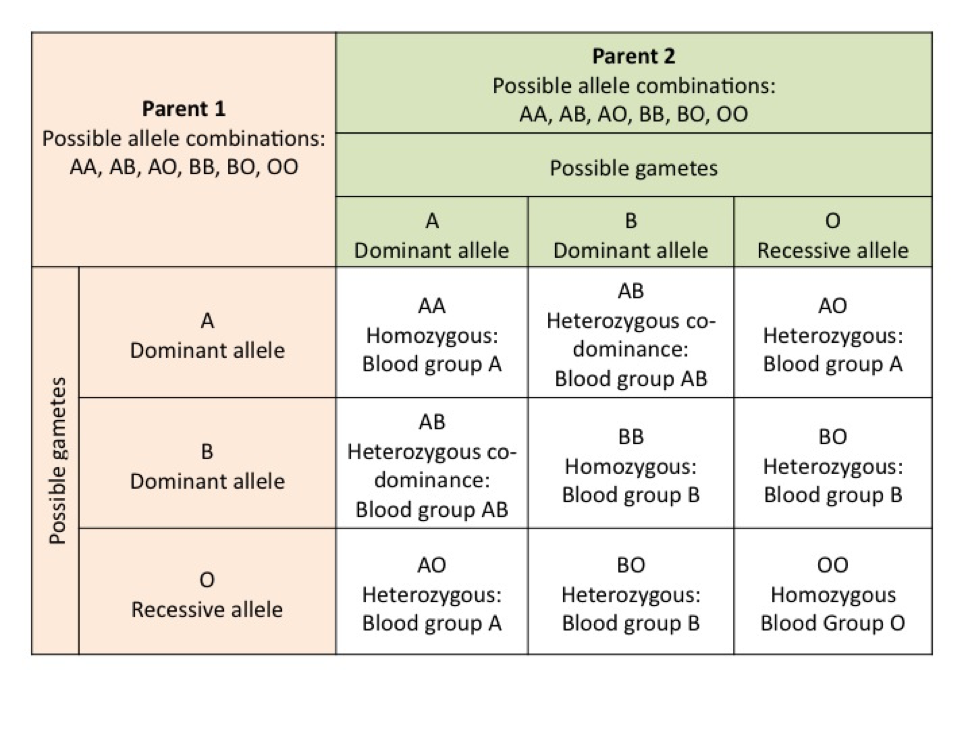
Punnett Square C for the probability of blood type inheritance, showing all possible combinations. Remember the parent will only have two alleles not all three
ABO blood grouping is an example of co-dominance. Here we have three genes involved. Two are dominant A and B, while the gene for the O blood group is recessive, that is, one must have two copies of that gene for it to be expressed (OO). A, B and O code for antigens on the surface of red blood cells. If the A antigen is present then you produce anti-B antibodies, and if the B antigen is present you produce anti-A antibodies. If one is blood group O no surface antigens are present and both anti-A and anti-B antibodies are produced. Antibody production is important for understanding blood transfusion reactions.
Punnet square C shows all the possible combinations for the ABO blood group system. A parent who is blood group A can have the genotype of either AA or AO, with the dominant allele A being expressed (and the one detected on a blood test). While one that is blood group AB expresses both the A and the B allele and antigens to both A and B are present. This individual does not produce antibodies to either A or B (see the cardiovascular module – the section on blood) and the alleles are co-dominant. That is, both alleles are expressed. The ABO blood typing system is only one of many blood typing systems. Another you would be familiar with is Rhesus negative or positive. This means you either have the Rhesus factor or you do not have it.
Punnet square C shows all the possible combinations for the ABO blood group system. A parent who is blood group A can have the genotype of either AA or AO, with the dominant allele A being expressed (and the one detected on a blood test). While one that is blood group AB expresses both the A and the B allele and antigens to both A and B are present. This individual does not produce antibodies to either A or B (see the cardiovascular module – the section on blood) and the alleles are co-dominant. That is, both alleles are expressed. The ABO blood typing system is only one of many blood typing systems. Another you would be familiar with is Rhesus negative or positive. This means you either have the Rhesus factor or you do not have it.
Sex-linked inheritance
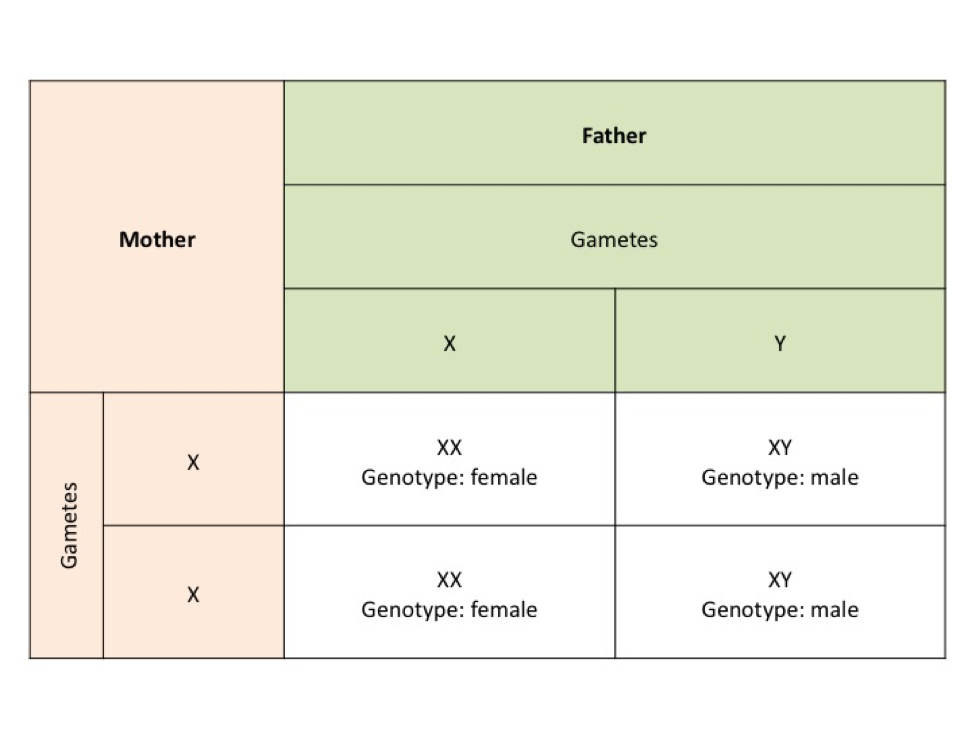
Punnet square D shows sex determination in offspring. Note the male always receives the X chromosome from the mother
Sex-linked inheritance is slightly different to autosomal inheritance in that a male only has one X chromosome which he inherits from the mother. The X and Y chromosome do not form a homologous pair. The Y is much smaller than the X chromosome and carries different genes to the X chromosome. This means that any allele on the X chromosome regardless of whether it is a recessive or dominant allele is expressed in the male. Females having two X chromosomes, one from the mother and one from the father, means that inheritance of traits for the female is similar to autosomal inheritance.
The male or female genotype is determined by whether a person has a Y chromosome. The Y chromosome holds the SRY gene that when switched on results in the production of testosterone. Testosterone is necessary for the development of male sex characteristics (see punnet square D). If the gene is not switched on, for example when not present in the female, or if the cell receptors for testosterone are resistant to the hormone testosterone then the male sex characteristics do not eventuate.
An example of a sex-linked trait that is often seen in men but rarely in women is baldness. Baldness is a recessive trait, and if the allele the male offspring receives from his mother carries the recessive gene then the male will eventually become bald. The female needs to carry two recessive alleles, one on each X chromosome in order to express this trait.
There are numerous sex-linked traits, and you will notice males are more commonly seen with these traits than women. We call these traits X-linked because it is the X chromosome that determines these phenotypic traits. The gene for red-green colour blindness is X-linked and is recessive; a person must have two copies of the gene to inherit colour blindness if they are female. The male however, only needs his X chromosome to carry the recessive gene to inherit the condition. A number of diseases are X-linked and include Duchenne muscular dystrophy and haemophilia.
The male or female genotype is determined by whether a person has a Y chromosome. The Y chromosome holds the SRY gene that when switched on results in the production of testosterone. Testosterone is necessary for the development of male sex characteristics (see punnet square D). If the gene is not switched on, for example when not present in the female, or if the cell receptors for testosterone are resistant to the hormone testosterone then the male sex characteristics do not eventuate.
An example of a sex-linked trait that is often seen in men but rarely in women is baldness. Baldness is a recessive trait, and if the allele the male offspring receives from his mother carries the recessive gene then the male will eventually become bald. The female needs to carry two recessive alleles, one on each X chromosome in order to express this trait.
There are numerous sex-linked traits, and you will notice males are more commonly seen with these traits than women. We call these traits X-linked because it is the X chromosome that determines these phenotypic traits. The gene for red-green colour blindness is X-linked and is recessive; a person must have two copies of the gene to inherit colour blindness if they are female. The male however, only needs his X chromosome to carry the recessive gene to inherit the condition. A number of diseases are X-linked and include Duchenne muscular dystrophy and haemophilia.
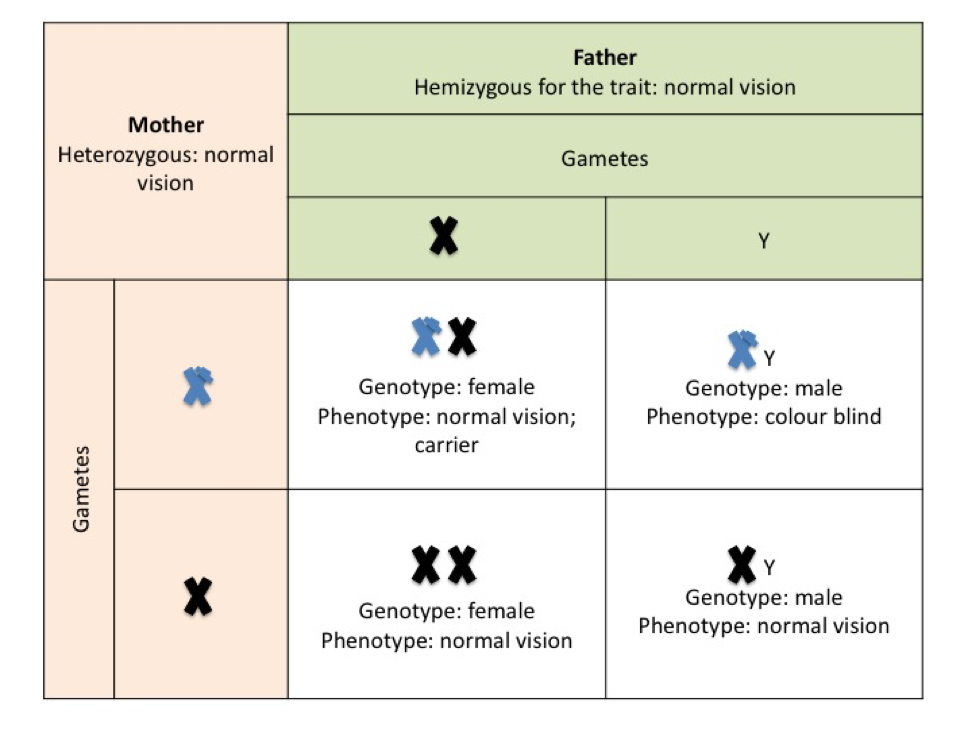
Punnet square E. Sex-linked inheritance for colour blindness, where the female is a carrier. Affected X chromosomes (recessive allele for the colour blind trait) are shown in blue in these examples.
In punnet square E the mother is a carrier of the recessive gene and the father has normal vision, all female offspring will have normal vision but there is a 50 % chance of a male offspring being colour blind.
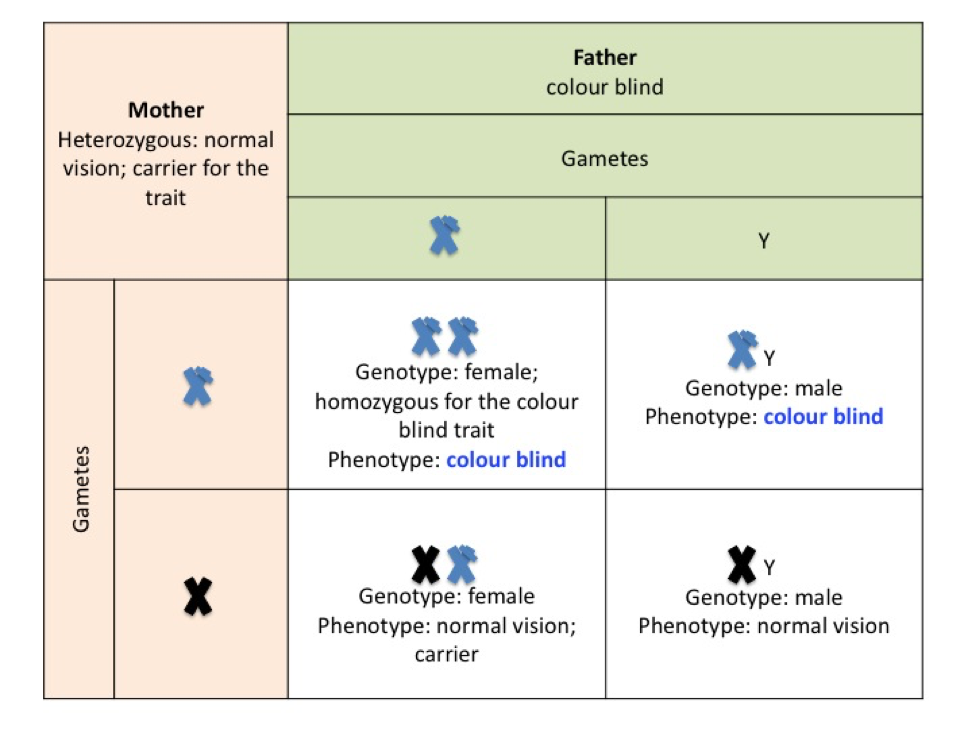
Punnet square F. Inheritance of colour blindness where the mother is a carrier and the father is colour blind
In punnet square F, the father is colour blind so has the mutated allele on his X chromosome – he will pass this X chromosome to his daughters. If the daughters inherit the mother’s affected chromosome as well they will be colour blind. If they receive the normal X chromosome from their mother they will be carriers of the faulty allele inherited from dad. Because sons inherit only the Y chromosome from their father, there is a one in two chance that any son will inherit the affected X chromosome from their mother and be colour blind.
Where the father is colour blind and the mother possesses the recessive gene, there is a 50 % chance of any offspring being colour blind.
Where the father is colour blind and the mother possesses the recessive gene, there is a 50 % chance of any offspring being colour blind.
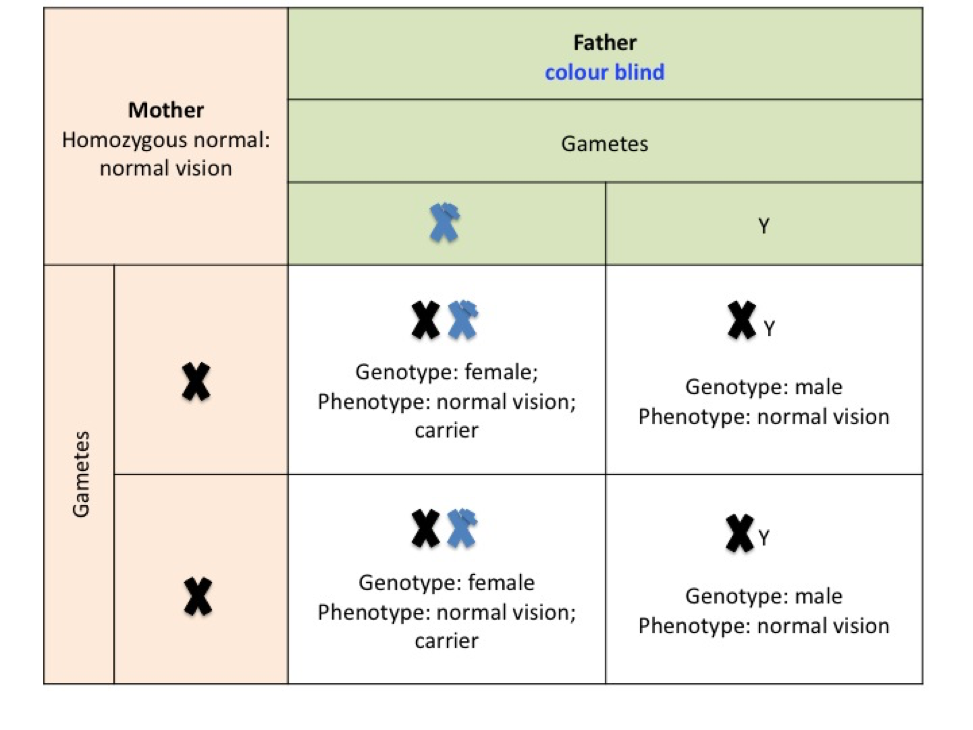
Punnet square G. Sex-linked inheritance of colour blindness, where the mother is homozygous dominant and the father is colour blind
If the mother does not carry the allele for colour blindness (punnet square G) then none of the male offspring will be colour blind. In this example the father is colour blind and all female offspring will inherit the recessive gene from him so that all girls will be carriers of the gene for colour blindness.
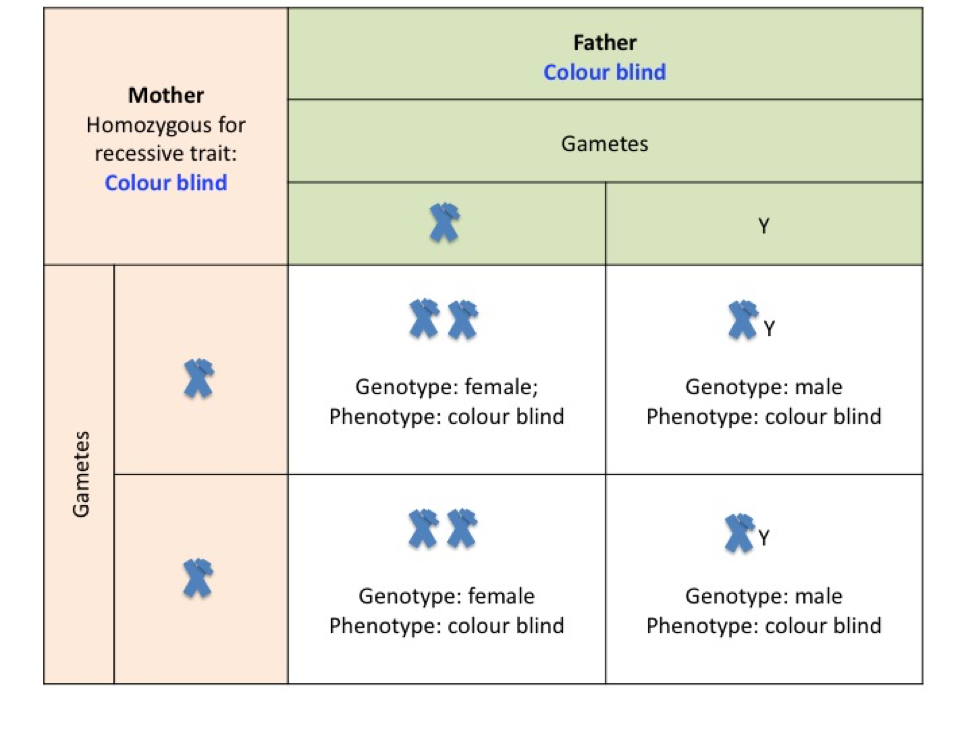
Punnet square H. Sex-linked inheritance for colour blindness where both parents are colour blind.
If both parents are colour blind then all offspring will be colour blind (see punnet square H)
Multifactorial traits
Some traits are influenced by more than one gene allele pair and are known as polygenic traits. An example is skin colour. Several genes regulate the production of the types of melanin by melanocytes. Production increases too in response to increased ultraviolet light exposure from the sun and just how well this process occurs is related to the genes you carry. Melanosomes in melanocytes produce the pigments eumelanin (brown tones) and pheomelanin (pink/red tones) which are then transferred to keratinocytes (skin cells). Another factor influencing skin tone is the pigment haemoglobin residing in our red blood cells which is of course heightened when we exercise as the blood moves towards the body surface to limit overheating. These factors, in combination, result in the many and varied shades of skin tone we see in the world.
Bibliography
Marieb, N. M. & Hoehn, K. (2013). Human Anatomy and Physiology. Ninth Edition. Person International Edition. Pearson Education Inc. USA.
Saladin, K. S. (2015). Anatomy and Physiology. The Unity of Form and Function. Seventh Edition. McGraw-Hill Education. USA.
Saladin, K. S. (2015). Anatomy and Physiology. The Unity of Form and Function. Seventh Edition. McGraw-Hill Education. USA.

