Learn more about the Lymphatic System & Immunity
In this module, you will learn more about
- The lymphatic system
- The immune system
- The stages of wound healing
Learn even more: See Tissues, Cardiovascular and Respiratory modules
1 The lymphatic system
The role of the lymphatic system is to return fluids that have leaked-out of the vascular system, back to the blood. It consists of a one-way network of vessels, lymph, lymph nodes and organs.
The lymph nodes and organs form part of the immune system. Fluid, called lymph, moving through lymph vessels pass through the lymph nodes where immune cells reside and screen the lymph for foreign particles. If foreign particles are present in the lymph they will activate immune defences.
Within the small intestine are specialised lymph vessels known as lacteals. Their role is to transport lipids.
The lymph nodes and organs form part of the immune system. Fluid, called lymph, moving through lymph vessels pass through the lymph nodes where immune cells reside and screen the lymph for foreign particles. If foreign particles are present in the lymph they will activate immune defences.
Within the small intestine are specialised lymph vessels known as lacteals. Their role is to transport lipids.
Lymph and Lymphatic Vessels
Lymph is a clear, low protein, colourless fluid, which varies in its constituents from place to place and time to time. For example, following a meal it may have a high lipid content. Lymph may also contain macrophages, lymphocytes from the lymph nodes, cellular debris, hormones; and foreign and endogenous particles such as bacteria, viruses and cancer cells.
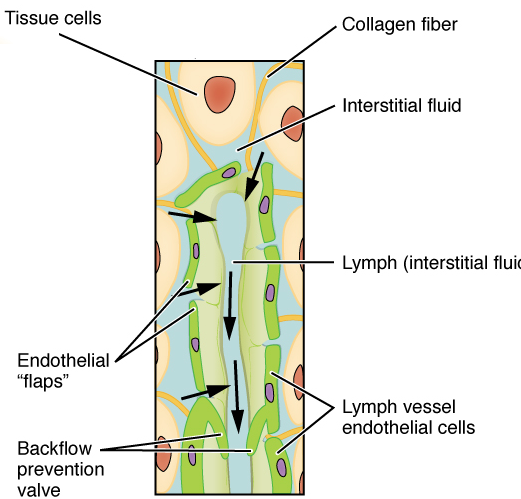
https://commons.wikimedia.org/wiki/File:2202_Lymphatic_Capillaries_small.png
Lymphatic vessels are similar to blood vessels but unlike the continuous structure of the circulatory system, they are closed at one end. Lymph moves in one direction only, and that is towards the heart. These vessels are closely associated with blood capillaries, but lymphatic endothelial cells, overlap to form valves. As valves, these overlapping endothelial cells can be pushed apart and allow fluid to enter the vessels (when tissue fluid pressure is high) and then close (when the tissue fluid becomes low). Because of the size of these valves, large particles such as: cell debris, large foreign particle such as bacteria, and immune cells; can easily pass into lymphatic vessels along with tissue fluid.
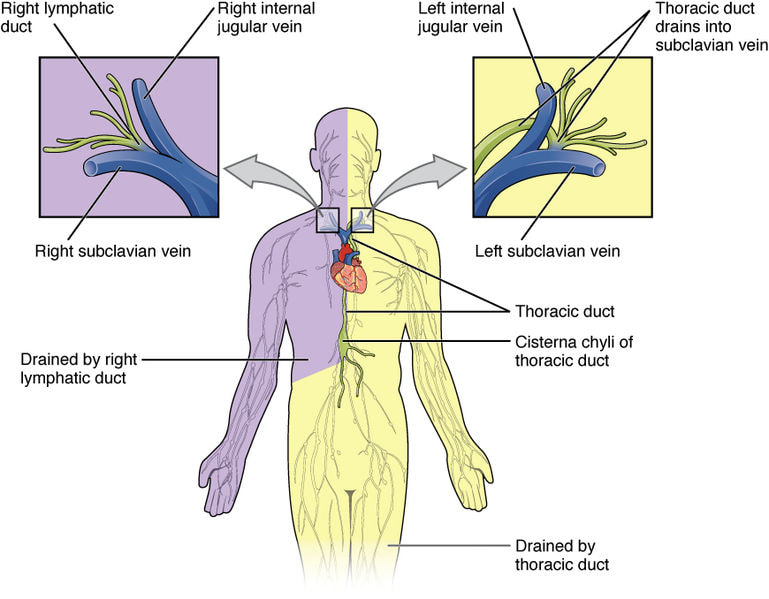
https://commons.wikimedia.org/wiki/File:2203_Lymphatic_Trunks_and_Ducts_System.jpg
The lymphatic vessels become larger as they move towards the heart ultimately emptying into the subclavian veins where lymph re-enters the circulatory system. Like veins lymphatic vessels contain one-way valves that prevent backflow of lymph. There is no pump, so pressure within the system is low with the main force being rhythmic contractions of the vessels as fluid stretches them. Just as with veins: skeletal muscle contraction, the pulsation of arteries in close contact with the lymphatic vessels, and the thoracic pump also drive lymph flow.
Lymphoid Cells and Tissue
Lymphatic tissues range from those found in mucous membranes (where the cells are loosely scattered) to that of lymphoid organs which are clearly defined and cell populations are dense. The cells found within lymphatic tissue have various roles in defence and immunity and include Natural Killer (NK) cells, T lymphocytes (“T cells”), B lymphocytes (“B cells”), macrophages, dendritic cells and reticular cells. These will be discussed further when looking at the immune system.
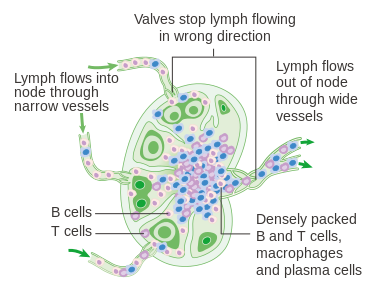
https://commons.wikimedia.org/wiki/File:Diagram_of_a_lymph_node_CRUK_022.svg
Lymph nodes are by far the most numerous of lymphoid organs and function to filter the lymph, and as sites for the activation of T- and B-lymphocytes. As the lymph drains through the nodes, about 99 % of debris and foreign particles found in the lymph are removed by macrophages and reticular cells.
This also means that foreign materials from different parts of the body are brought to sites with abundant immune cells (lymphocytes, macrophages and dendritic cells). Exposure to foreign materials will activate cells of the immune system. In the case of the lymphocytes of the lymph nodes these activated cells can migrate into the blood stream and onto the site where the foreign material entered the body. In this sense we describe one of the roles of the lymphatic system as immunosurveillance.
This also means that foreign materials from different parts of the body are brought to sites with abundant immune cells (lymphocytes, macrophages and dendritic cells). Exposure to foreign materials will activate cells of the immune system. In the case of the lymphocytes of the lymph nodes these activated cells can migrate into the blood stream and onto the site where the foreign material entered the body. In this sense we describe one of the roles of the lymphatic system as immunosurveillance.
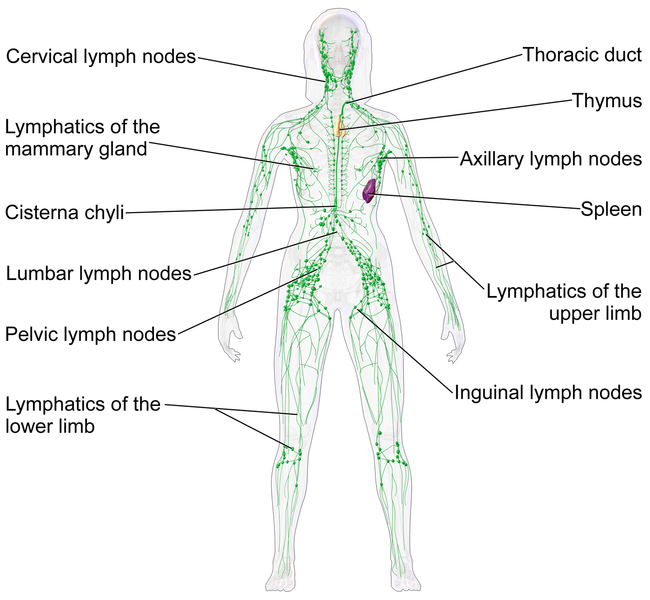
https://commons.wikimedia.org/wiki/File:Blausen_0623_LymphaticSystem_Female.png
While lymph nodes are found throughout the body they are more highly concentrated in some parts of the body namely: the neck (cervical) the armpits (axillary) the thorax, abdomen, intestinal and mesenteric regions, the groin (inguinal) and the back of the knee (popliteal). You would become aware of them when you have an infection, as lymph nodes draining the infected region of the body will become swollen and painful.
Patches of lymphoid tissue are found in the respiratory tract (the tonsils) and the small intestine (Peyer’s patches). These sites are in contact with the external environment, so like the lymph nodes these masses of lymphoid tissue are also involved in immunosurveillance.
The spleen is the largest of the lymphoid organs. It is encapsulated and lies within the abdominal cavity just below the diaphragm. The spleen consists of red pulp and white pulp, the former being the site where non-functional erythrocytes and blood pathogens are destroyed. The white pulp forms part of the immune system and is composed mainly of lymphocytes. While the spleen is a site of both immune surveillance, and proliferation of lymphocytes, it has several other functions:
The lymphatic system serves quite a different additional role.
Lipids aren’t very soluble in water-based fluids like the plasma. The lipids molecules that are absorbed in the cells of the intestinal mucosa are combined with specific proteins to form lipoprotein particles which is a major way lipid is carried in the body fluids. The lipoprotein particles produced in the mucosal cells are of a particular structure and composition and are called chylomicrons. These particles are quite big; but they are readily taken up into the lymphatics in the intestinal sub-mucosa. This means that amongst the different things transported in lymph are the fats from the food we eat.
Patches of lymphoid tissue are found in the respiratory tract (the tonsils) and the small intestine (Peyer’s patches). These sites are in contact with the external environment, so like the lymph nodes these masses of lymphoid tissue are also involved in immunosurveillance.
The spleen is the largest of the lymphoid organs. It is encapsulated and lies within the abdominal cavity just below the diaphragm. The spleen consists of red pulp and white pulp, the former being the site where non-functional erythrocytes and blood pathogens are destroyed. The white pulp forms part of the immune system and is composed mainly of lymphocytes. While the spleen is a site of both immune surveillance, and proliferation of lymphocytes, it has several other functions:
- Removal of blood debris which include defective blood cells and platelets.
- Splenic macrophages remove debris and foreign particles through the process of phagocytosis.
- Storage of the breakdown products of erythrocytes for recycling.
- Storage of platelets, and monocytes (circulating immune cells), for future use.
The lymphatic system serves quite a different additional role.
Lipids aren’t very soluble in water-based fluids like the plasma. The lipids molecules that are absorbed in the cells of the intestinal mucosa are combined with specific proteins to form lipoprotein particles which is a major way lipid is carried in the body fluids. The lipoprotein particles produced in the mucosal cells are of a particular structure and composition and are called chylomicrons. These particles are quite big; but they are readily taken up into the lymphatics in the intestinal sub-mucosa. This means that amongst the different things transported in lymph are the fats from the food we eat.
2 The immune system
Our bodies encounter numerous foreign materials daily: bacteria, fungi, viruses and also non-infectious materials like pollen and toxins; and yet for the most part we remain healthy and do not succumb to disease. This is due to our immune system. The major cells of the immune system are the white blood cells or leukocytes, which constantly circulate between the blood and the tissues and whose role is to destroy (or neutralise or just promote removal of) substances that the body recognises as not part of us which could potentially cause us harm. Immunologists distinguish the bits of the body that are us as “self” and anything foreign as “non-self”.
White blood cells/Leukocytes
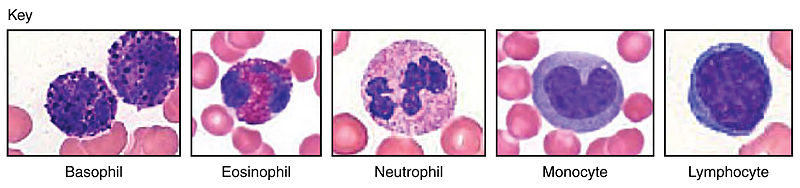
https://commons.wikimedia.org/wiki/File:1916_Leukocyte_Key.jpg
Identification of leukocytes is based on their colour following staining and their shape of the respective nuclei There are several different types of leukocytes and they all derive from stem cells in the red bone marrow.
Granulocytes (because they have prominent granules (“little grains”):
Mononuclear Cells (because their nuclei are not segmented like granulocytes and so it really does look they have a single nucleus)
The immune system is conventionally divided into the innate immune system and the adaptive immune system, although these two arms of our defence system work together to deal with foreign materials but their relative contribution will vary depending on the foreign object.
The following is a basic introduction to the immune system. If you wish to learn more about it then a good immunology text book or scientific journal articles should be sought where you can see the developments that are taking place in this evolving field. Before we move on to the cells and protein of the immune system we need to consider the first line of defence to invasion provided by other parts of the body.
Granulocytes (because they have prominent granules (“little grains”):
- neutrophils
- eosinophils
- basophils and the tissue residing mast cells
Mononuclear Cells (because their nuclei are not segmented like granulocytes and so it really does look they have a single nucleus)
- monocytes/macrophages
- lymphocytes: including natural killer cells, T- and B lymphocytes.
The immune system is conventionally divided into the innate immune system and the adaptive immune system, although these two arms of our defence system work together to deal with foreign materials but their relative contribution will vary depending on the foreign object.
The following is a basic introduction to the immune system. If you wish to learn more about it then a good immunology text book or scientific journal articles should be sought where you can see the developments that are taking place in this evolving field. Before we move on to the cells and protein of the immune system we need to consider the first line of defence to invasion provided by other parts of the body.
Barriers to invasion
- The skin: As a multi-layered tissue (with the upper layers being made up largely of a protein that many micro-organisms can’t degrade) the skin is a physical barrier to invasion. Added to this much of the skin is dry with a low pH and so “hostile” to microbial colonisation. The parts that aren’t (where there is body hair) support commensal populations of microorganism which will essentially starve out or even kill any invading microorganism that arrive
- Sweat and sebum: Sweat is secreted onto the skin to promote body cooling, it’s saltiness means it is also anti-microbial. Sebum is an oily secretion which serves a moisturising function for the skin and hair but is also anti-microbial
- Mucous membranes: These line the tracts of the body which have openings to the external environment. Those openings being: the mouth, the nostrils, the anus, the urethra, and (additionally in females) the vagina. Mucus is secreted upon the inner lining of each of the tracts and inhibits the capacity of micro-organisms to adhere. Also the mucus will contain anti-microbial agents. Acid secretion is a feature of gastric and vaginal lining which is also anti-microbial.
There are two interacting arms to the immune system:
- the innate
- the adaptive
The Innate Immune System
The innate immune system is made up of: phagocytic cells, natural killer (NK), cells, and antimicrobial proteins. The response to the invading pathogen, often resullts in inflammatory responses and fever.
The distinguishing features of the innate system are:
The distinguishing features of the innate system are:
- It is rapid that is it begins within seconds of the invasion
- It is non-specific. It relies on the identification of molecules on (or released or triggered by) the invading pathogen that are shared by different pathogens. These molecules are recognised by receptors on the cells of the immune system
- It does not become stronger or faster or more efficient if the same pathogen re-infects you. The response is “hard-wired” in your genes, that is it’s innate
Cells of the innate immune system
Phagocytes
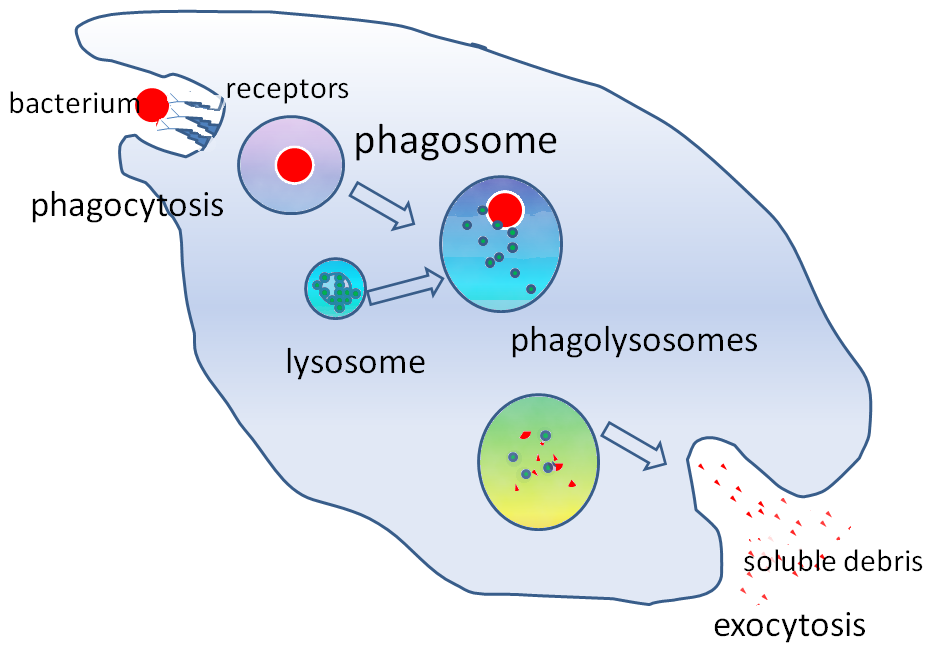
The process of phagocytosis https://upload.wikimedia.org/wikipedia/commons/e/e6/Phagocytosis2.png
|
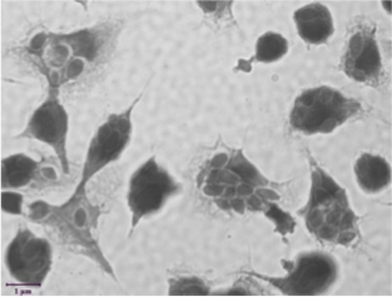
Macrophages phagocytosing yeast © J. Salvage-Jones, 2016
|
Phagocytosis is the process by which the phagocyte binds and engulfs a foreign body via cytoplasmic extensions. The phagocytic cell uses lysosomal enzymes to destroy the enclosed foreign matter. As well as directly binding to the microorganism, the phagocyte will recognise and engulf microorganisms literally coated in proteins of the immune system either proteins of the complement cascade or proteins produced by the adaptive immune system (antibodies).
There are two types of phagocytes:
In general neutrophils are the first to arrive on the scene. In the act of dealing with invasion neutrophils die. These dead neutrophils form a major component of pus.
In contrast, macrophages are long-lived. Several parts of the body have a resident macrophage population because of the chronic exposure to microorganisms for example: the liver (“Kupffer cells” and the alveoli (mast cells).
What brings the phagocytes from out of the blood stream and into the invaded tissue? At sites of invasion immune cells in the tissues and the cells that line the local blood vessels release chemotactic agents and also the cells lining the blood vessels insert adhesive proteins on the surface facing the blood. The combined result is that the white blood cells first adhere to the lining of the blood vessel, then cross the vessel wall and then migrate towards the site of invasion.
Macrophages serve additional purposes. They are important in wound healing through the removal of dead and damaged cells. They also have a central role in the regulation of the adaptive immune system.
There are two types of phagocytes:
- neutrophils
- cells that are derived from monocytes called macrophages.
In general neutrophils are the first to arrive on the scene. In the act of dealing with invasion neutrophils die. These dead neutrophils form a major component of pus.
In contrast, macrophages are long-lived. Several parts of the body have a resident macrophage population because of the chronic exposure to microorganisms for example: the liver (“Kupffer cells” and the alveoli (mast cells).
What brings the phagocytes from out of the blood stream and into the invaded tissue? At sites of invasion immune cells in the tissues and the cells that line the local blood vessels release chemotactic agents and also the cells lining the blood vessels insert adhesive proteins on the surface facing the blood. The combined result is that the white blood cells first adhere to the lining of the blood vessel, then cross the vessel wall and then migrate towards the site of invasion.
Macrophages serve additional purposes. They are important in wound healing through the removal of dead and damaged cells. They also have a central role in the regulation of the adaptive immune system.
Natural Killer (NK) Cells
These cells are large lymphocytes and are not phagocytic. As agents of the innate immune system, they target, and can recognise and destroy a wide range of abnormal cells including tumor cells, and virally-infected cells. They also regulate the activity of cells of both the innate and adaptive immune systems.
Antimicrobial Proteins
There are two classes known as:
Interferons are secreted by leukocytes when they are infected with viruses. This alerts other cells within the vicinity of the infection and protects them from becoming infected. Interferons bind to cell surface receptors and induce the release of antiviral proteins which either breakdown the virus or prevent it from replicating. Furthermore, these proteins activate macrophages and NK cells, leading to the destruction of the infected cell before more viral particles are released.
Complement is a large group of proteins that circulate in the blood in inactive forms. In a process like that of coagulation: they are aligned in a sequence where activation of one member brings about the activation of the next one in the sequence.
- interferons and
- complement.
Interferons are secreted by leukocytes when they are infected with viruses. This alerts other cells within the vicinity of the infection and protects them from becoming infected. Interferons bind to cell surface receptors and induce the release of antiviral proteins which either breakdown the virus or prevent it from replicating. Furthermore, these proteins activate macrophages and NK cells, leading to the destruction of the infected cell before more viral particles are released.
Complement is a large group of proteins that circulate in the blood in inactive forms. In a process like that of coagulation: they are aligned in a sequence where activation of one member brings about the activation of the next one in the sequence.
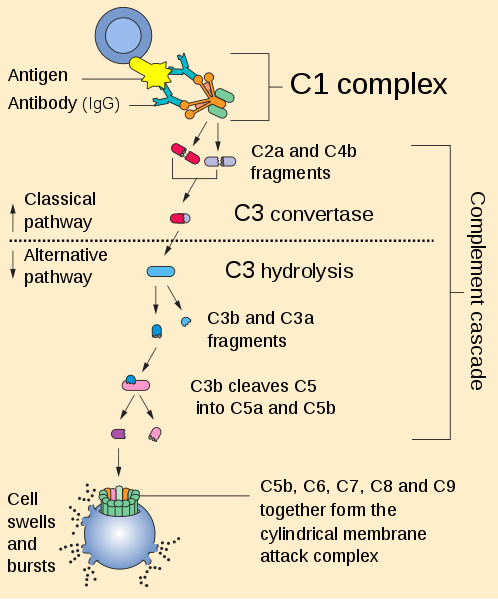
https://commons.wikimedia.org/wiki/File:Complement_pathway.svg
The diagram illustrates that:
- There is a “cascade” of reactions initiated once an invading organism is present
- The system is involved in both the innate (“Alternative pathway”) and adaptive (“Classical pathway) immune responses
- In either case the cascade of reactions includes inactive complement proteins being cleaved to yield active fragments
- Activated complement components form complexes most notably: the Membrane Attack Complex which literally punches holes in the plasma membrane of the invader
- What is not shown is that fragments have other roles in innate response
The Inflammatory Response
The inflammatory response is activated whenever the tissues of the body are injured by any agent.
Its purpose is to:
As nurses you will be able to identify the five cardinal signs of inflammation:
Its purpose is to:
- recruit the cells of the innate (and later the adaptive) immune response to the site of damage
- prevent the spread of damage
- destroy pathogens
- remove any host cell debris and prepare the region for wound healing
As nurses you will be able to identify the five cardinal signs of inflammation:
- Redness
- Heat
- Swelling
- Pain
- Loss of function
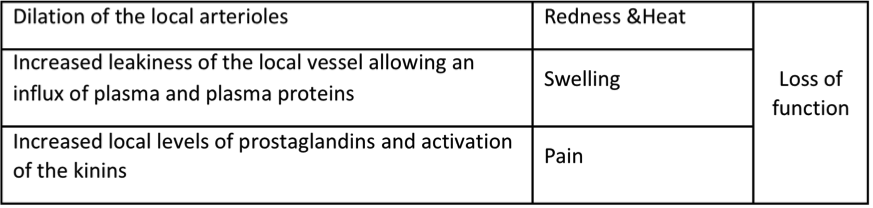
© Dr David van Reyk 2018: Manifestations of what happens at an inflamed site
|
Central to the inflammatory response are the mast cells. They leave the bone marrow as immature cells and migrate into vascularised tissues where they mature. They can be found in many tissues, particularly those that have contact with the external world such as the skin, airways and intestines. They are structurally similar to basophils in that their cytoplasm is filled with granules. Triggers associated with tissue injury will cause the mast cells to exocytose the contents of their granules. Key components of these are histamine and serotonin which bring about the changes in the local vessels that characterise the inflammatory response. Notably though in the airways and GIT histamine causes constriction of these tracts. Mast cells are also the source of prostaglandins and a related group of molecules called the leukotrienes. Some mast cell products are chemotactic. Mast cell also have a role in adaptive immune system which is particularly signficant when that role contributes to allergy and anaphylaxis.
|
https://upload.wikimedia.org/wikipedia/commons/5/5d/Blausen_0077_Basophil.png
|
Fever
Body temperature is very tightly controlled around a set point and under normal circumstances it can fluctuate around the set point over a narrow range of roughly 36.5–37.5 degrees Celsius. When inflammation occurs, soluble factors known as cytokines can drive a shift upward in the set point so that the body’s temperature will rise. This is what we call fever.
The function of fever is not fully understood but could include:
Fever should not be seen as a natural process allowed to “run its course”. This is because some people may experience serious ill effects from the fever. Also body temperatures of 43 degrees Celsius and above can be fatal. There are other safer means to promote the body’s immune responses such as bed rest while the person is ill.
The function of fever is not fully understood but could include:
- as a sign to the person that they have an infection
- impeding the multiplication and function of invading microorganisms
- keeping the body at a temperature where parts of the immune system work at their best
Fever should not be seen as a natural process allowed to “run its course”. This is because some people may experience serious ill effects from the fever. Also body temperatures of 43 degrees Celsius and above can be fatal. There are other safer means to promote the body’s immune responses such as bed rest while the person is ill.
Cytokines
Think of these as locally acting hormones. That is, like hormones they are produced and released by particular cells, most notably (but not only) cell of the immune system. Like hormones they will target cells that have the specific receptor for that particular cytokine. Like hormones cytokines can stimulate cells or inhibit their function.
Two distinct functions of cytokines are
There are many cytokines including the different interleukins, interferon-gamma and tumour necrosis factor-alpha
Two distinct functions of cytokines are
- some act upon the cells that produce and release them;
- they are released into the local tissue although there will be “leakage” into general circulation.
There are many cytokines including the different interleukins, interferon-gamma and tumour necrosis factor-alpha
Other granulocytes
- Eosinophils have central roles in allergy and dealing with parasite infections.
- Basophils have roles which overlap with those of mast cells
The Adaptive Immune System
The adaptive immune system is made up of:
The distinguishing features of the adaptive system are:
Central to your understanding of the adaptive immune system are two concepts:
- antigen-presenting cells
- T- and B-lymphocytes
- antibodies
The distinguishing features of the adaptive system are:
- It is delayed as it can take several days to be optimal. This relates to the fact that involves a large increase in the lymphocyte population
- It is specific. In the whole population of T- and B-lymphocytes, individual “members” will only “recognise” and respond to a specific invading pathogen, particle or molecule
- It becomes stronger, faster and more efficient if the same pathogen re-infects you. The response is adaptive. The specific mix of lymphocytes in a person is essentially a history of her exposure to particular pathogens and other foreign materials
Central to your understanding of the adaptive immune system are two concepts:
- Antigen recognition
- Immunological memory
Antigens
An antigen is any molecule that activates an immune response. They can be free such as in the case of toxins, or may make up part of the plasma membrane or cell wall of a pathogen. They are usually complex molecules that are not found naturally occurring within the human body. Only part of the antigen needs to be antigenic (stimulate an immune response), and that region is known as an antigenic determinant or epitope. Most antigens have more than one epitope, and as such can stimulate different members of the body’s lymphocyte population.
Antigen Presenting Cells (APC)
APC include macrophages, B-lymphocytes and dendritic cells. Examples of the latter will be found, for example, residing in the skin and lymph nodes. APC will engulf foreign material and then present fragments of their antigens on their cell surface for T-lymphocytes to recognise. For a T-cells to recognise the fragment, it must be presented in combination with particular members of the major histocompatibility complex (MHC) family of proteins.
The Major Histocompatibility Complex (MHC) Family of Proteins
The MHC is a family of proteins of which classes I and II are of interest here. Class II are only found on APC. These are combined with antigen fragments from foreign cells that the APC has engulfed. Class I are present on nearly all body cells and will combine with protein fragments that come from inside the cell. These intracellular protein fragments are usually bits of digested self-proteins but also can be fragments of a virus if it has infected a cell. Without being complexed to an antigen fragment, MHC molecules will not trigger an immune response.
B-lymphocytes and humoural immunity
|
|
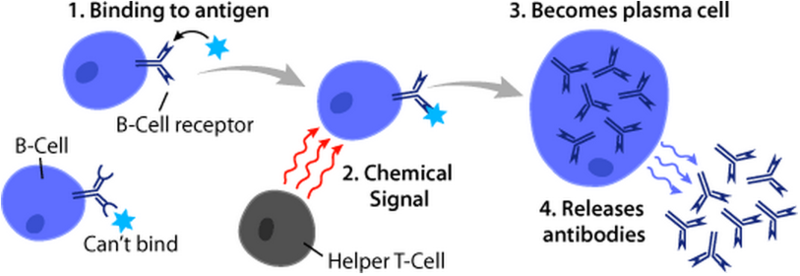
https://commons.wikimedia.org/wiki/File:B_cell_function.png
|
Humoural immunity refers to antibody-mediated immunity and the central player in humoural immunity is the B lymphocyte.
Recall the whole lymphocyte population consists of an army of specialists. Each member will only recognise and respond to a particular epitope of a particular antigen. This is because on its surface are receptors for that, and only that, particular epitope. When any particular B-lymphocyte encounters the epitope it will recognise it becomes activated. The key features of this being
Recall the whole lymphocyte population consists of an army of specialists. Each member will only recognise and respond to a particular epitope of a particular antigen. This is because on its surface are receptors for that, and only that, particular epitope. When any particular B-lymphocyte encounters the epitope it will recognise it becomes activated. The key features of this being
- The B-lymphocyte will undergo cell division: it will proliferate
- The daughter cells differentiated to what are called plasma cells
- The plasma cells are the source of specialised immune proteins called antibodies. The antibodies that any particular plasma cell produces will only recognise and bind to the same epitopes that triggered the formation of that particular plasma cell
Antibodies and humoural immunity
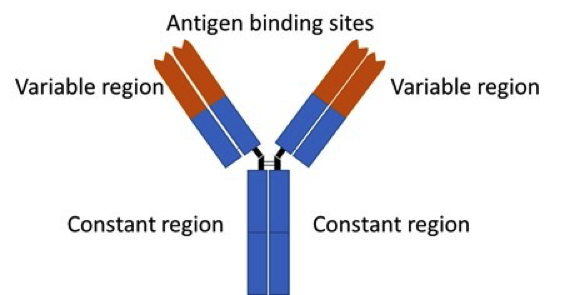
Simplistic representation of the structure of antibodies
As the name suggests the variable region will vary in structure and amino acid composition from one antibody to the next. This is the part of the antibody which will recognise and bind to a particular epitope. The constant region is the same for antibodies of the same class (more on this later).
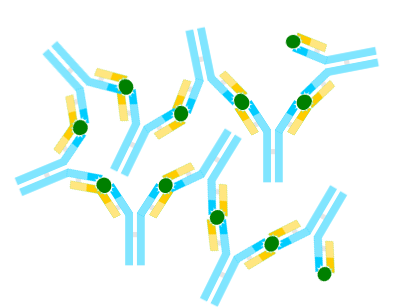
https://upload.wikimedia.org/wikipedia/commons/a/a1/Antigen-antibody-complex.png
Several antibody molecules may bind individual epitopes on several epitope bearing structures to form what is called an antibody-antigen complex. The diagram is not drawn to scale. The (blue) antibodies and the (green) epitope are invariably much smaller than the (pale orange) epitope-bearing molecule or cell.
Antibodies serve a number of functions:
While there are numerous antibodies, each is specific for a specific antigen, they are grouped into five classes known as IgA, IgD, IgE, IgG and IgM. Each class of antibody has a slightly different structure and function. For example
Plasma cells can switch between antibodies that they produce, although regardless of class, the antibodies will retaindspecificity for the same antigen. Class switching is particularly important when it comes to the switch from a primary to a secondary immune response which we will go through later.
Antibodies serve a number of functions:
- Neutralisation involves the antibody binding to and blocking specific sites on viruses or toxin preventing them from binding to the host cells. Anti-venoms given to people who get snake and spider bites work this way
- Opsonisation involves coating a foreign cell in specific antibodies, this aids greatly in the phagocytosis of the invader
- Removal is where antibody-antigen complexes are engulfed by phagocytic cells.
- Complement fixation results when several antibody molecules bind to the same cell, and this can trigger activation of the complement cascade: the so-called “Classical pathway” of complement activation
While there are numerous antibodies, each is specific for a specific antigen, they are grouped into five classes known as IgA, IgD, IgE, IgG and IgM. Each class of antibody has a slightly different structure and function. For example
- IgE play a central role in allergy
- IgA are involved in immune responses at mucous membranes
- IgD serve as the receptors on the surface of B-lymphocytes
Plasma cells can switch between antibodies that they produce, although regardless of class, the antibodies will retaindspecificity for the same antigen. Class switching is particularly important when it comes to the switch from a primary to a secondary immune response which we will go through later.
T-lymphocytes and cell-mediated immunity
T-lymphocytes directly interact with their target cell. However, just like B-lymphocytes when it recognises its particular antigen, that particular T-lymphocyte will differentiate and proliferate.
There are several populations of T-lymphocytes but this module will focus on just two. They are named after a characteristic molecule present on their surface:
There are several populations of T-lymphocytes but this module will focus on just two. They are named after a characteristic molecule present on their surface:
- CD4 or ”Helper” T-lymphocytes
- CD8 or ”Cytotoxic” T-lymphocytes
|
Helper T-lymphocytes are activated by a specific antigen fragment (and this will vary from one particular lymphocyte to the next) combined MHC class II molecules on the surface of an APC. They get the name “Helper” because help to activate other immune cells such as B-lymphocytes, macrophages, dendritic cells and other T-lymphocytes. The diagram illustrates how this would work with a B-lymphocyte. (You can also see a Helper cell “in action” in the earlier diagram about B-lymphocytes becoming plasma cells). How Helper cell helps is through the release of cytokines which turn regulate the function of other cells of the immune system.
|
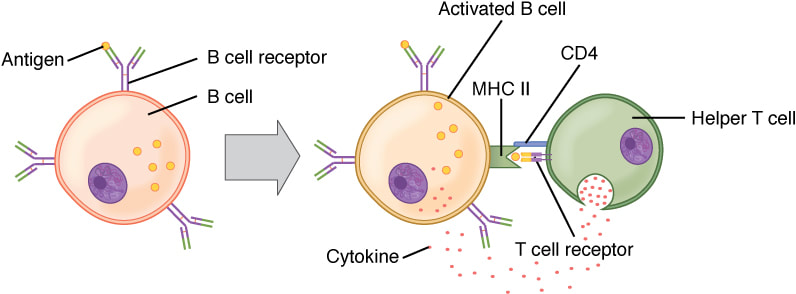
https://commons.wikimedia.org/wiki/File:2224_T_and_B_Cell_Binding.jpg
|
|
CD8 T-lymphocytes cytotoxic in that they target infected cells and destroy them. These are the only cells of the adaptive immune system that directly targets infected cells by binding to antigen presented by MHC I molecules. Because MHC I molecules are found on nearly all body cells, cytotoxic T-lymphocytes survey their environment for any cell displaying foreign antigens. Their main target is virus infected cells, but they will also attack cells that are infected with bacteria or parasites, and cancer cells. As the diagram indicates, cytotoxic T-lymphocytes can do their work through secreting proteins (granzymes and perforins) that cause the infected cell to lyse. Alternatively, they can initiate a process called apoptosis which is a highly regulated and controlled way for a cell to die
|
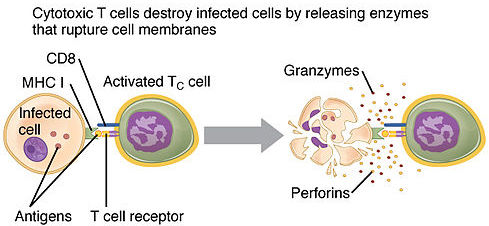
https://commons.wikimedia.org/wiki/File:2219_Pathogen_Presentation.jpg
|
Immunological Memory: Primary and Secondary Immune Responses
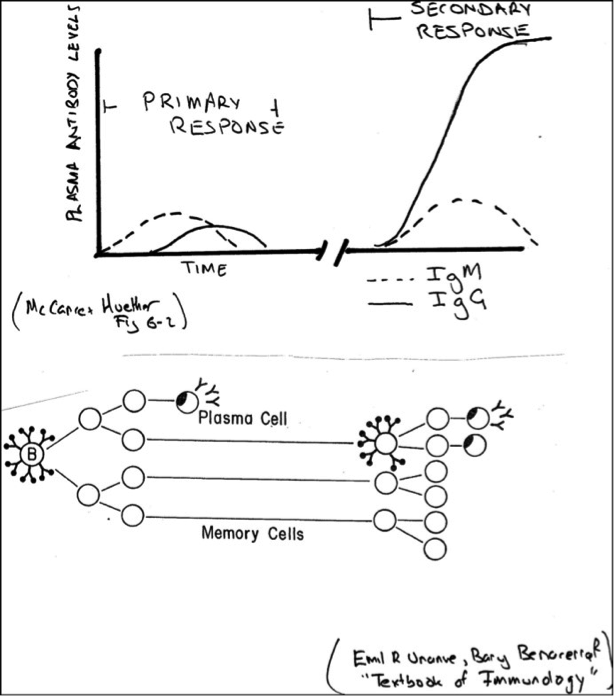
When a B- or T-lymphocyte becomes activated it proliferates. What that would mean is that during any particular immune response the lymphocytes which recognise the particular antigenic trigger will substantially outnumber the lymphocytes which don’t. Once the source of antigen is dealt with most of the lymphocytes die through apoptosis. Most but not all. In fact, there is now a persistent bias in the number of those particular lymphocytes.
Think of it this way if I was exposed to tetanus toxin and you weren’t; then if we measured how many tetanus toxin-responsive lymphocytes we both have, I would have more
That is my immune response has adapted to the fact that I was once exposed to tetanus toxin. This is what we mean by immunological memory. You can appreciate the next time I am exposed to tetanus toxin I will have a bigger response because I have tetanus toxin-specific memory cells. In the bottom part of the diagram: there was just a few (represented by one) B-lymphocytes before antigen exposure. Antigen exposure lead to proliferation. Some those daughter cells stuck around as memory cells. When the same antigen reappeared there were more cells ready to go.
It’s not just a numbers game. That is illustrated in the top part of the diagram. When it comes to humoural responses, the first exposure involves production of mainly IgM class antibodies: a primary immune response. With the second exposure there is a switch to the more effective IgG class. You will also see the antibody levels are much higher and production occurs earlier. These are all features of secondary immune response.
Think of it this way if I was exposed to tetanus toxin and you weren’t; then if we measured how many tetanus toxin-responsive lymphocytes we both have, I would have more
That is my immune response has adapted to the fact that I was once exposed to tetanus toxin. This is what we mean by immunological memory. You can appreciate the next time I am exposed to tetanus toxin I will have a bigger response because I have tetanus toxin-specific memory cells. In the bottom part of the diagram: there was just a few (represented by one) B-lymphocytes before antigen exposure. Antigen exposure lead to proliferation. Some those daughter cells stuck around as memory cells. When the same antigen reappeared there were more cells ready to go.
It’s not just a numbers game. That is illustrated in the top part of the diagram. When it comes to humoural responses, the first exposure involves production of mainly IgM class antibodies: a primary immune response. With the second exposure there is a switch to the more effective IgG class. You will also see the antibody levels are much higher and production occurs earlier. These are all features of secondary immune response.
Clinical considerations of the Adaptive Immune Response
The two central features of the adaptive immune response: specificity and memory; have two very important clinical applications
- The first is diagnostic. Recall that a particular antigen will only activate particular B-lymphocytes which will produce unique antibodies. Unique because they will only bind to epitopes of that antigen. In the case of infectious diseases, you can detect the exposure to a pathogen even if it is difficult to detect the pathogen itself. Instead you look for a high level of antibodies to that pathogen is the person’s blood (“elevated antibody titre”)
- The second is therapeutic. If you had a really nasty infectious agent, you could perhaps isolate antigenic parts of it (or develop a safe version of it) and inject that into a person. This will elicit a primary response without causing the disease in the person AND with the subsequent production of memory cells. Can you guess what will happen when that person encounters the actual pathogen subsequently? A more effective and faster secondary immune response will occur and hopefully the response will be strong enough that the person does not succumb to the disease. The process that has been just described of “priming” the immune response with a non-hazardous form of the pathogen of interest is the basis of vaccination.
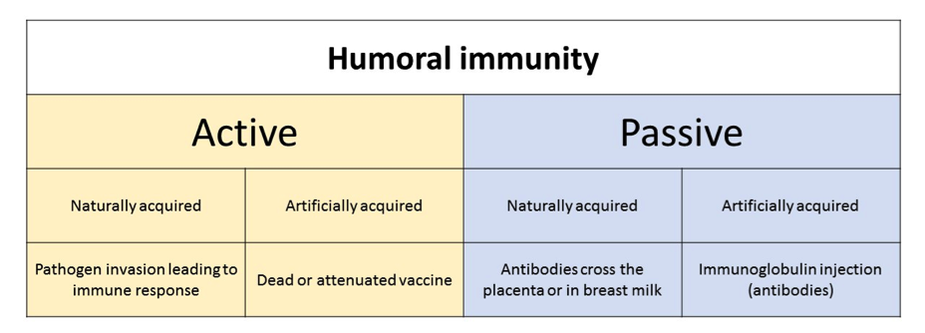
Vaccines and vaccinations
Vaccination is the administration of a vaccine to produce immunity from particular disease. Vaccines act to stimulate the body’s immune system with a weakened (attenuated) or killed form of the causative microorganism. Examples of diseases for which vaccines are available in Australia include:
- Polio
- Rotavirus
- Pertussis
- Tetanus
- Influenza
- Hepatitis A & B
3 The stages of wound healing
Wound healing includes two major components:
- regeneration - the replacement of lost cells and tissues with cells of the same type
- repair - the more common type of healing and usually results in scar formation.
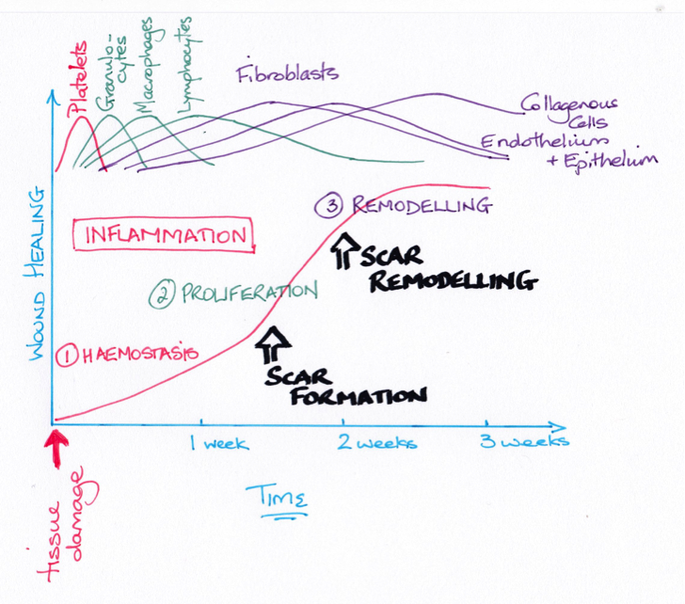
© Dr Patricia Logan 2017: Timeline for healing
When a wound occurs, the first reaction of the human body is to maintain stability through physiological inflammatory responses which attack, confine, and kill microorganisms.
The inflammatory response is a sequential reaction to cell injury neutralising and diluting inflammatory agents, removing necrotic materials, and establishing a suitable environment for wound healing and repair.
The inflammatory response is a sequential reaction to cell injury neutralising and diluting inflammatory agents, removing necrotic materials, and establishing a suitable environment for wound healing and repair.
Inflammation is divided into three basic types.
The inflammatory response is divided into overlapping and interdependent phases:
- Acute inflammation, where the healing occurs in 2–3 weeks and usually leaves no residual damage
- Subacute inflammation has the acute process features, however, it lasts longer
- Chronic inflammation which can last for weeks, months, or sometimes years
The inflammatory response is divided into overlapping and interdependent phases:
- the vascular response
- cellular response
- inflammatory exudate formation
- healing
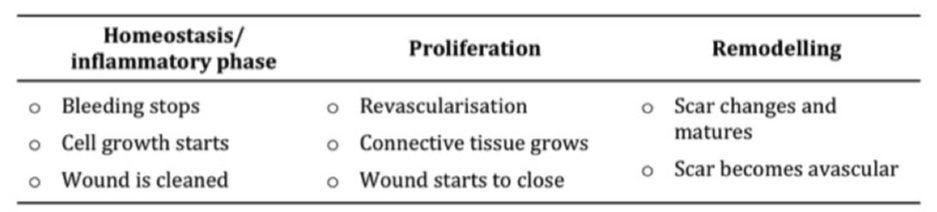
Homeostasis/inflammatory phase
Haemostasis means 'stopping the bleeding'. During the vascular inflammation phase a number of processes occur simultaneously. Platelets release vasoconstrictive substances reducing blood loss. While vasodilatation and increased hydrostatic pressure increases blood supply (hyperaemia) to the injured area. Increased capillary permeability allows leakage of fluids into interstitial spaces to dilute the toxins.
Growth factors are also secreted by platelets and macrophages. While platelet-activating factor (PAF) induces platelet aggregation (clotting). Epidermal growth factor (EGF) stimulates keratinocyte (skin cells) proliferation and migration of cells which stabilises epidermal turnover resulting in barrier function.
Growth factors are also secreted by platelets and macrophages. While platelet-activating factor (PAF) induces platelet aggregation (clotting). Epidermal growth factor (EGF) stimulates keratinocyte (skin cells) proliferation and migration of cells which stabilises epidermal turnover resulting in barrier function.
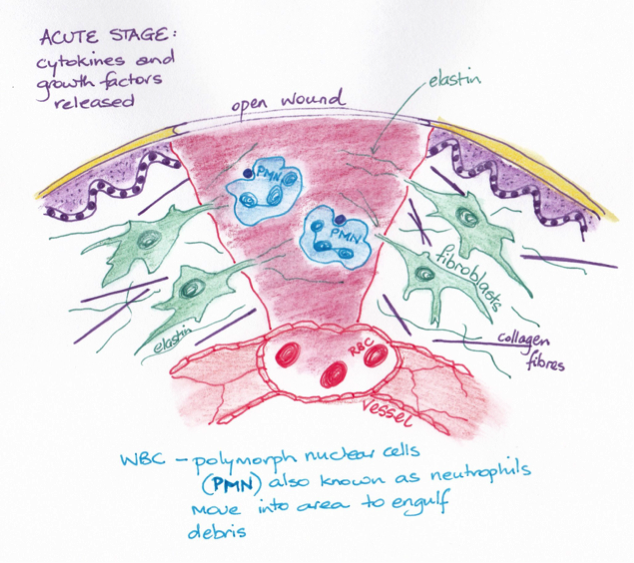
© Dr Patricia Logan 2017: Acute stage
During the cellular phase, white blood cells respond to the injury. Chemotactic factors attract white blood cells to the wound, where they adhere (margination) and migrate (emigration) through the vessel wall. Histamines are released from mast cells within hours after injury. Prostaglandin is also released assisting to regulate platelet aggregation, control inflammation, and vascular permeability. Phagocytes attracted to wound enzymatically degrade foreign matter and dead tissue, forming exudate (wound drainage).
Inflammatory exudate consists of fluid and leucocytes, which move from the circulation to the wound. The exudate transports leukocytes, antibodies, dilute toxins and irritating substances as well as transporting nutrients for tissue repair. The nature and quantity of exudate is dependent upon the type and severity of wound and the tissues involved. Types of inflammatory exudate include:
Inflammatory exudate does not indicate infection during the inflammatory phase, as this is the body’s physiological response to injury. Systemic clinical manifestations include fever and leukocytosis (increase in WBC count due to any cause). Pyrogens released from injured cells and white blood cells stimulate the hypothalamus’ thermo-regulating centre increasing temperature, which helps to fight any infection. Leukocytosis is initiated when damaged cells release leukopoietin and stimulate white blood cell and neutrophil production. With continued inflammation, immature neutrophils are produced, reflected as “bands” on the WBC differential count (also called a “shift to the left”).
Finally, fibroblasts secrete fibrin and wall off the area.
In a clean acute wound, the inflammatory phase lasts approximately 3 to 5 days. However, a wound complicated by infection or necrosis, will prolong this phase. As the inflammatory phase prepares the site for new tissue growth and wound closure by cleaning the wound; suppression of the inflammatory phase delays wound healing. Inflammatory phase suppression can be caused by: radiation therapy, malnutrition, dehydration, chronic disease processes (e.g. diabetes mellitus, chronic obstructive pulmonary disease), or medications (e.g. corticosteroids).
Inflammatory exudate consists of fluid and leucocytes, which move from the circulation to the wound. The exudate transports leukocytes, antibodies, dilute toxins and irritating substances as well as transporting nutrients for tissue repair. The nature and quantity of exudate is dependent upon the type and severity of wound and the tissues involved. Types of inflammatory exudate include:
- Serous exudate: watery and low in protein
- Serosanguineous drainage: has serous and some blood
- Fibrinous exudate: thick, sticky, and high in protein
- Purulent exudate: infective organisms, leukocytes, and cellular debris
- Haemorrhagic exudate: contains red blood cells.
Inflammatory exudate does not indicate infection during the inflammatory phase, as this is the body’s physiological response to injury. Systemic clinical manifestations include fever and leukocytosis (increase in WBC count due to any cause). Pyrogens released from injured cells and white blood cells stimulate the hypothalamus’ thermo-regulating centre increasing temperature, which helps to fight any infection. Leukocytosis is initiated when damaged cells release leukopoietin and stimulate white blood cell and neutrophil production. With continued inflammation, immature neutrophils are produced, reflected as “bands” on the WBC differential count (also called a “shift to the left”).
Finally, fibroblasts secrete fibrin and wall off the area.
In a clean acute wound, the inflammatory phase lasts approximately 3 to 5 days. However, a wound complicated by infection or necrosis, will prolong this phase. As the inflammatory phase prepares the site for new tissue growth and wound closure by cleaning the wound; suppression of the inflammatory phase delays wound healing. Inflammatory phase suppression can be caused by: radiation therapy, malnutrition, dehydration, chronic disease processes (e.g. diabetes mellitus, chronic obstructive pulmonary disease), or medications (e.g. corticosteroids).
Proliferation phase
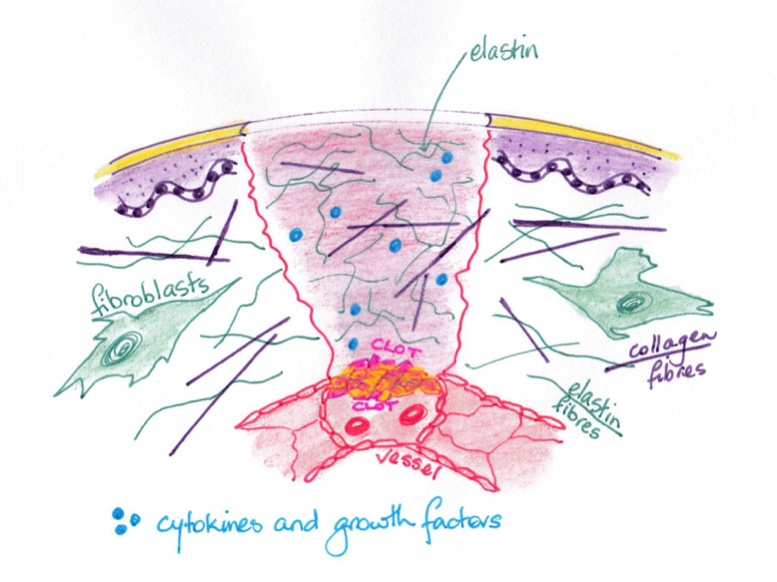
© Dr Patricia Logan 2017: Proliferation stage
Reconstruction occurs in the proliferation phase, which lasts up to 3 weeks in a healthy person. Growth factors stimulate vascular buds formation and vascular loops regrowth. Additionally, endothelial cells regenerate and when stimulated by angiogenic substances form new capillaries. If sufficient blood circulation is sufficient this begins within 3 days of injury. Also fibroblasts begin to produce connective tissue at the wound margins. Fibroblasts also produce collagen fibres; in collagen synthesis, fibrin proteins form a matrix to support the newly forming tissue. Myofibroblasts develop at wound edges and induce wound contraction.
Granulation tissue containing newly formed capillaries, proliferating fibroblasts, and residual inflammatory cells develops approximately 3 days after the initial injury. At this stage the wound is beefy red and moist. If granulation tissue is slow to form, it may be due to:
o Insufficient blood supply
o Wound Infection
o Lack of one of the necessary growth factors for revascularization
During epithelialization the wound starts to close from its margins, and cells migrate from the wound edges over the granulating tissue; covering the wound with a layer of very fragile new pink or pearl-like skin. A moist wound bed that is well perfused with oxygenated blood is required to support wound healing.
Granulation tissue containing newly formed capillaries, proliferating fibroblasts, and residual inflammatory cells develops approximately 3 days after the initial injury. At this stage the wound is beefy red and moist. If granulation tissue is slow to form, it may be due to:
o Insufficient blood supply
o Wound Infection
o Lack of one of the necessary growth factors for revascularization
During epithelialization the wound starts to close from its margins, and cells migrate from the wound edges over the granulating tissue; covering the wound with a layer of very fragile new pink or pearl-like skin. A moist wound bed that is well perfused with oxygenated blood is required to support wound healing.
Remodelling Phase
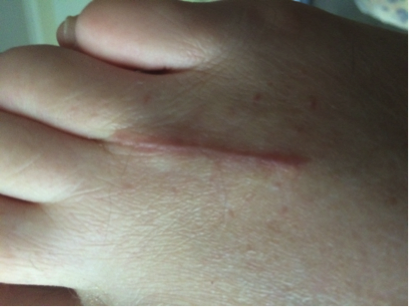
At 1 month: Scar tissue remodels over time becoming avascular and so white in colour
|
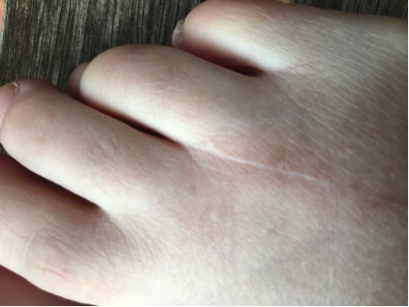
At 14 months: Scar tissue remodels over time becoming avascular and so white in colour
|
Remodelling occurs after the wound is closed; approximately 3 weeks after the injury. Closure can however still be in process from 6 months to 2 years later. The closer together that the edges might be brought together, the smaller the amount of scar tissue formed and the faster the wound closes. As the scar changes and matures; its bulk decreases and colour changes from pink to pearly white. The incisional strength increases causing contraction which can cause collagen fibres to become taut. Finally, the scar tissue shrinks, leaving visible tension line. It is important to remember healed wounds achieve only 65–80% of their original tensile strength, so they are at a greater risk of future breakdown.
You will learn more about the inflammatory responses and stages of wound healing during your pre-registration course. This knowledge is vital to promote effective wound healing and management.
You will learn more about the inflammatory responses and stages of wound healing during your pre-registration course. This knowledge is vital to promote effective wound healing and management.
Bibliography
Bunker, N. & Fagan, A. (2015). Nursing care of people with vascular and lymphatic disorders. In P. LeMone, K. Burke, T. Levett-Jones, T. Dwyer, L. Moxham, ….. D. Raymond. (Eds.). Medical surgical nursing: Critical thinking for person-centred care (2nd Aust. ed.). Frenchs Forest, Australia: Pearson.
Copstead, L-E., & Banasik, J. (Eds.). (2013). Pathophysiology. (5th ed.). St Louis, MO:Elsevier
Gordon, C. & Craft, J. (2015). The structure and function of the cardiovascular and lymphatic system. In J. Craft, C. Gordon & A. Tiziani (Eds.), Understanding pathophysiology (2nd Aust. ed.). (pp. 558-605). Chatswood, Australia: Mosby.
Marieb, E.N. & Hoehn, K. (2014). Human Anatomy & Physiology, (9th ed.). St Louis, MO:Elsevier
Wagner, K. D. & Hardin-Pierce, M. G. (2014). High-acuity nursing (6th ed.). Upper Saddle River, NJ:Pearson
Marieb, E. N. & Hoehn,K. (2013). Human Anatomy and Physiology. Pearson Education, USA. (9th Ed).
Mckinley, M. P., O’Loughlin, V. D. & Bidle, T. S. (2015). Anatomy and Physiology; An Integrative Approach. McGraw-Hill Education, Europe. (2nd Ed).
Saladin. K. S. (2015). Anatomy and Physiology; The Unity of Form and Function. McGraw-Hill Education, USA. (7th Ed).
Copstead, L-E., & Banasik, J. (Eds.). (2013). Pathophysiology. (5th ed.). St Louis, MO:Elsevier
Gordon, C. & Craft, J. (2015). The structure and function of the cardiovascular and lymphatic system. In J. Craft, C. Gordon & A. Tiziani (Eds.), Understanding pathophysiology (2nd Aust. ed.). (pp. 558-605). Chatswood, Australia: Mosby.
Marieb, E.N. & Hoehn, K. (2014). Human Anatomy & Physiology, (9th ed.). St Louis, MO:Elsevier
Wagner, K. D. & Hardin-Pierce, M. G. (2014). High-acuity nursing (6th ed.). Upper Saddle River, NJ:Pearson
Marieb, E. N. & Hoehn,K. (2013). Human Anatomy and Physiology. Pearson Education, USA. (9th Ed).
Mckinley, M. P., O’Loughlin, V. D. & Bidle, T. S. (2015). Anatomy and Physiology; An Integrative Approach. McGraw-Hill Education, Europe. (2nd Ed).
Saladin. K. S. (2015). Anatomy and Physiology; The Unity of Form and Function. McGraw-Hill Education, USA. (7th Ed).

