Learn more about the Gastrointestinal system
In this module, you will learn more about
- The gastronintestinal system overall
- Regulation of gastric function
- The GIT and digestion
- Metabolism
- Metabolic states
- Important roles of the liver
Learn even more: see Chemistry, Cells, Tissues and Homeostasis modules
1 The gastrointestinal system overall
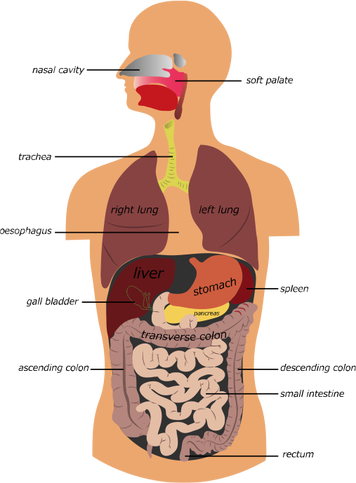
The gastrointestinal tract and accessory organs (Depositphotos.com)
The gastrointestinal tract (GIT) is a long muscular tube that runs from the mouth and ends at the anus. Ingested food is progressively broken-down by chemicals; enzymes, hormones, and acid, and absorbed into the blood. The structure of the tract is dedicated to its specific function with different organs contributing to the digestive process.
Organs of the digestive tract include:
Organs of the digestive tract include:
- the mouth
- pharynx
- oesophagus
- stomach
- small intestine (the duodenum, the jejunum, and the ileum)
- large intestine
- accessory organs - teeth, tongue, salivary glands, liver, gall bladder and pancreas.
Digestive process
There are six stages to digestion:
Terminology Note:
Enzyme names usually end in –ase. There are exceptions to this rule such as pepsin and trypsin.
‘Entero’ as a part of any word indicates a component of the gastric tract eg. enterocyte refers to cells of the gastric tract.
- Ingestion is basically the intake of food.
- Propulsion which moves food along the digestive tract. It starts with swallowing (deglutition) followed by peristalsis. Peristalsis is a series of repeated contractions and relaxation of the muscular walls of the tract which moves food along the tract.
- Mechanical breakdown of food products breaks them into smaller pieces which them more accessible to enzymes. Chewing and the muscular contractions of the stomach mash up ingested food.
- Digestion relies on the secretion of enzymes from the GIT that breakdown complex molecules into simpler substances that can be used by the body.
- Absorption involves the uptake of nutrients from the lumen of the GIT into the blood or lymph.
- Defecation is the process by which unused and undigested components are excreted from the body.
Terminology Note:
Enzyme names usually end in –ase. There are exceptions to this rule such as pepsin and trypsin.
‘Entero’ as a part of any word indicates a component of the gastric tract eg. enterocyte refers to cells of the gastric tract.
2 Regulation of gastric function
Both the nervous and endocrine systems are involved in the control of gastric secretions.
They work together to either increase gastric secretions or to suppress them.
There are three phases to gastric secretion and as their names suggest, stimuli act on the head, the stomach and the small intestine:
They work together to either increase gastric secretions or to suppress them.
There are three phases to gastric secretion and as their names suggest, stimuli act on the head, the stomach and the small intestine:
1 The cephalic phase
This starts with a response to the thought, sight, smell or taste of food and lasts only a few minutes.
Sensory input is sent to the hypothalamus, which in turn signals to the medulla oblongata. Impulses are sent down the vagus nerve to parasympathetic enteric ganglia, neurons of which stimulate the glands of the stomach, parietal cells and G cells to secrete HCl and gastrin.
Sensory input is sent to the hypothalamus, which in turn signals to the medulla oblongata. Impulses are sent down the vagus nerve to parasympathetic enteric ganglia, neurons of which stimulate the glands of the stomach, parietal cells and G cells to secrete HCl and gastrin.
2 The gastric phase
This is initiated when food reaches the stomach and lasts for 3-4 hours.
Ingested food stimulates this phase in three ways; distension of the stomach wall and a rising the pH (making the environment less acidic), and the presence proteins which also act as buffers that bind hydrogen (H+) ions.
Distension of the stomach wall activates reflexes that lead to the release of acetylcholine (ACh) which stimulates the release of gastric juice.
Partially digested proteins act a chemical stimuli and the rising pH directly activates enteroendocrine cells (G cells) to secrete gastrin.
Gastrin and ACh as well as histamine, act on the parietal cells to release HCl and the chief cells to secrete pepsinogen.
The break-down of protein into smaller peptides and amino acids stimulates G cells to produce more gastrin and this results in a positive feedback loop which accelerates protein digestion. As protein digestion progresses and the smaller peptides and amino acids leave the stomach the pH falls lower and lower until it drops below 2. At this point the high acidity inhibits the secretions from the parietal cells and the chief cells thus preventing the pH from falling further. This negative feedback loop helps to maintain an optimal environment for enzymic actions in the stomach.
Ingested food stimulates this phase in three ways; distension of the stomach wall and a rising the pH (making the environment less acidic), and the presence proteins which also act as buffers that bind hydrogen (H+) ions.
Distension of the stomach wall activates reflexes that lead to the release of acetylcholine (ACh) which stimulates the release of gastric juice.
Partially digested proteins act a chemical stimuli and the rising pH directly activates enteroendocrine cells (G cells) to secrete gastrin.
Gastrin and ACh as well as histamine, act on the parietal cells to release HCl and the chief cells to secrete pepsinogen.
The break-down of protein into smaller peptides and amino acids stimulates G cells to produce more gastrin and this results in a positive feedback loop which accelerates protein digestion. As protein digestion progresses and the smaller peptides and amino acids leave the stomach the pH falls lower and lower until it drops below 2. At this point the high acidity inhibits the secretions from the parietal cells and the chief cells thus preventing the pH from falling further. This negative feedback loop helps to maintain an optimal environment for enzymic actions in the stomach.
3 The intestinal phase
This sees the chyme arriving at the duodenum and gastric activity being moderated by hormones and nervous reflexes.
As food starts to enter the duodenum it starts to stretch.
Further distension of the duodenum with chyme that contains H+, partially digested proteins and semi-digested fats triggers the enterogastric reflex.
This reflex inhibits vagal stimulation of the stomach and stimulates sympathetic neurones which send inhibitory signals to the stomach causing the pyloric sphincter to constrict, stopping further entry of chyme into the duodenum.
Gastric activity is stopped, which protects the duodenum from the effects of excessive acidity and from chyme overload. During this phase enterogastrones are released. These are intestinal hormones which include secretin, cholecystokinin (CCK), and vasoactive intestinal peptide (VIP), which all inhibit gastric secretions and motility when the stomach is full. They also stimulate the gall bladder to contract which will be discussed shortly.
As food starts to enter the duodenum it starts to stretch.
Further distension of the duodenum with chyme that contains H+, partially digested proteins and semi-digested fats triggers the enterogastric reflex.
This reflex inhibits vagal stimulation of the stomach and stimulates sympathetic neurones which send inhibitory signals to the stomach causing the pyloric sphincter to constrict, stopping further entry of chyme into the duodenum.
Gastric activity is stopped, which protects the duodenum from the effects of excessive acidity and from chyme overload. During this phase enterogastrones are released. These are intestinal hormones which include secretin, cholecystokinin (CCK), and vasoactive intestinal peptide (VIP), which all inhibit gastric secretions and motility when the stomach is full. They also stimulate the gall bladder to contract which will be discussed shortly.
3 The GIT and digestion
The mouth
The mouth (oral cavity or buccal cavity), is lined with thick squamous epithelium and is kept moist by secretions from the salivary glands.
Its digestive functions include ingestion, taste, mastication (chewing), chemical digestion, and swallowing (deglutition).
Non-digestive functions include speech and respiration.
It is composed of:
From the mouth food enters the oropharynx and then the laryngopharynx, the mucosa of which contain friction-resistant stratified squamous epithelium. This epithelium is supplied with mucous producing glands. Two layers of skeletal muscle surround the pharynx; a deep layer of longitudinal and a superficial layer of circular constrictor muscles. The contraction of these superficial muscles and secretion of mucous into the pharynx allows the smooth passage of food towards the oesophagus.
The oesophagus is a straight muscular tube about 25 cm long lying inferior to the larynx and posterior to the trachea. It remains collapsed when empty. From the laryngopharynx, the oesophagus moves through the mediastinum and passes through the diaphragm at the oesophageal hiatus. After another 3-4 cm it meets the stomach at the cardiac orifice. When food reaches this point there is a slight pause before it enters the stomach, due to the constriction of the cardiac sphincter, a small band of circular smooth muscle. This muscle prevents food from being regurgitated and, along with mucous secreting cells surrounding the sphincter, protects the oesophagus from the corrosive effects of stomach acid.
Its digestive functions include ingestion, taste, mastication (chewing), chemical digestion, and swallowing (deglutition).
Non-digestive functions include speech and respiration.
It is composed of:
- The lips and cheeks that help to retain food within the oral cavity in order that it can be chewed.
- The teeth that masticate food, breaking it into smaller pieces. In adults, there are usually 16 teeth in the mandible and 16 in the maxilla. Each jaw contains four incisors, two canines, four premolars and up to six molars designed for their specific functions of: cutting; puncturing and shredding; and crushing, shredding and grinding, respectively.
- The tongue composed of bundles of skeletal muscle fibres that manipulates the food between the teeth, while also mixing the food with saliva to form a compact mass known as a bolus. The superior surface of the tongue contains filiform papillae which roughen the tongue’s surface and enables one to lick semisolid foodstuffs, and also provides surface friction for manipulating foods. Scattered across the surface of the tongue are fungiform papillae, and on its lateral surface are foliate papillae which house the taste buds.
- Saliva which moistens and cleanses the mouth, and binds food particles into a bolus whilst lubricating it with mucous for ease of swallowing. The salivary glands empty their contents into the mouth. Saliva contains mostly water (97-99.5 %) and is slightly acidic (pH ~ 6.75). It also contains salivary amylase for the breakdown of starch, and lingual lipase for lipid digestion. 1-1.5 L of saliva is produced per day. Most saliva is produced in response to food entering the oral cavity but the smell of food will also cause salivation. Mechanoreceptors and chemoreceptors located there send messages to the salivatory nuclei in the brain stem which increases parasympathetic nervous system (PNS) activity. Impulses are sent to the salivary glands via the facial and glossopharyngeal nerves, resulting in an increase in the output of saliva. What do you think happens to levels of saliva in response to strong sympathetic NS stimulation? (What happens to your mouth when you are anxious?).
From the mouth food enters the oropharynx and then the laryngopharynx, the mucosa of which contain friction-resistant stratified squamous epithelium. This epithelium is supplied with mucous producing glands. Two layers of skeletal muscle surround the pharynx; a deep layer of longitudinal and a superficial layer of circular constrictor muscles. The contraction of these superficial muscles and secretion of mucous into the pharynx allows the smooth passage of food towards the oesophagus.
The oesophagus is a straight muscular tube about 25 cm long lying inferior to the larynx and posterior to the trachea. It remains collapsed when empty. From the laryngopharynx, the oesophagus moves through the mediastinum and passes through the diaphragm at the oesophageal hiatus. After another 3-4 cm it meets the stomach at the cardiac orifice. When food reaches this point there is a slight pause before it enters the stomach, due to the constriction of the cardiac sphincter, a small band of circular smooth muscle. This muscle prevents food from being regurgitated and, along with mucous secreting cells surrounding the sphincter, protects the oesophagus from the corrosive effects of stomach acid.
The stomach
The stomach is a j-shaped muscular sac that functions primarily as a food storage organ.
It has an internal volume of about 50 ml when empty and 1-1.5 L when full.
The stomach lies in the upper left quadrant of the peritoneal cavity and is almost hidden by the liver and the diaphragm. It is divided into four regions:
This pyloric canal terminates in the pylorus which forms a narrow passage into the duodenum. Surrounding the pylorus is the pyloric sphincter which controls the passage of chyme from the stomach to the duodenum.
The stomach is innervated by both parasympathetic and sympathetic nerve fibres of the autonomic nervous system and its blood supply is via branches of the celiac trunk. All blood leaving the stomach and intestines enters the hepatic portal circulation before returning to the heart.
The stomach is involved in both mechanical and chemical digestion and its microscopic anatomy is specifically suited to these functions.
It has an internal volume of about 50 ml when empty and 1-1.5 L when full.
The stomach lies in the upper left quadrant of the peritoneal cavity and is almost hidden by the liver and the diaphragm. It is divided into four regions:
- The cardiac region or cardia surrounds the cardiac orifice.
- The fundus is a dome shaped region that bulges superolaterally to the cardia.
- The body or corpus is the largest part of the stomach and lies distal to the cardia.
- The pyloric region is a narrower pouch that lies at the inferior end of the stomach and is divided into the superior and wider pyloric antrum which narrows to form the pyloric canal.
This pyloric canal terminates in the pylorus which forms a narrow passage into the duodenum. Surrounding the pylorus is the pyloric sphincter which controls the passage of chyme from the stomach to the duodenum.
The stomach is innervated by both parasympathetic and sympathetic nerve fibres of the autonomic nervous system and its blood supply is via branches of the celiac trunk. All blood leaving the stomach and intestines enters the hepatic portal circulation before returning to the heart.
The stomach is involved in both mechanical and chemical digestion and its microscopic anatomy is specifically suited to these functions.
The muscularis externa of the stomach wall contains three layers, a longitudinal, a circular, and an inner layer of smooth muscle which runs obliquely.
This muscular arrangement allows the stomach to mix, churn and move food towards the pylorus. The mixing and churning action turns the food into the liquid called chyme.
The stomach mucosa is covered with simple columnar glandular epithelium. The apical regions of these cells contain mucin which when secreted mixes with water to become alkaline mucous. This mucous lines the stomach and traps a layer of bicarbonate, that, along with tight junctions that join the epithelial cells together, protects the stomach wall against the acid. The mucosa and submucosa contain longitudinal wrinkles (rugae) when empty. As the stomach fills these rugae flatten-out giving the mucosa a smooth appearance. Dotted throughout the mucosa are gastric pits which lead to gastric glands that secrete gastric juice.
Cells found in the gastric glands include:
This muscular arrangement allows the stomach to mix, churn and move food towards the pylorus. The mixing and churning action turns the food into the liquid called chyme.
The stomach mucosa is covered with simple columnar glandular epithelium. The apical regions of these cells contain mucin which when secreted mixes with water to become alkaline mucous. This mucous lines the stomach and traps a layer of bicarbonate, that, along with tight junctions that join the epithelial cells together, protects the stomach wall against the acid. The mucosa and submucosa contain longitudinal wrinkles (rugae) when empty. As the stomach fills these rugae flatten-out giving the mucosa a smooth appearance. Dotted throughout the mucosa are gastric pits which lead to gastric glands that secrete gastric juice.
Cells found in the gastric glands include:
- Parietal cells that secrete hydrochloric acid (HCl) and intrinsic factor. HCl makes the stomach contents very acidic (pH 1-1.5), which enables the conversion of pepsinogen to pepsin, and is then necessary for the activation of pepsin. This acidity also helps in the digestive process by denaturing proteins and breaking down the cell walls of plant matter, while also destroying bacteria ingested with the food. Intrinsic factor is required for vitamin B12 absorption within the small intestine.
- Chief cells that produce pepsinogen and secrete lipases for the digestion of lipids.
- Enteroendocrine cells release hormones such as histamine, serotonin, somatostatin and gastrin, which regulates stomach secretions and motility.
The small intestine
The small intestine is a coiled tube filling most of the abdominal cavity.
It runs from the pyloric sphincter to the ileocecal sphincter where it joins to the large intestine.
It is divided into three parts:
It runs from the pyloric sphincter to the ileocecal sphincter where it joins to the large intestine.
It is divided into three parts:
- the duodenum - approx. 25 cm long
- the jejunum - approx. 1-1.7m long, and
- the ileum -approx.1.6-2.7m long.
The duodenum is the first section of the small intestine and lies retroperitoneally, inferior to the liver and the stomach, and arcing around the head of the pancreas. The bile duct and the pancreatic duct join together to deliver bile and pancreatic juice to the lumen of the duodenum. These fluids mix with the acidic chyme that has been delivered from the stomach into the duodenum.
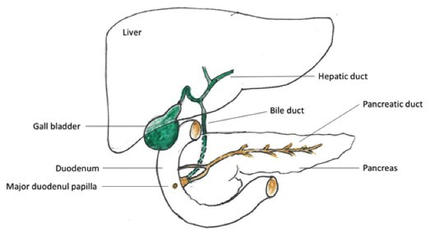
Accessory organs of the gastrointestinal tract.
The jejunum mainly occupies the umbilical region and has a thick and muscular wall with a rich blood supply. Most digestion and absorption takes place here.
The ileum lies within the hypogastric region and has a thinner, less muscular and less vascular wall. Lymphatic nodules known as Peyer patches are found attached to the ileum, becoming larger as the ileum approaches the large intestine. The ileum joins the cecum of the large intestine at the ileocecal junction, where a muscular sphincter, the ileocecal valve, is found. This valve controls the passage of food into the large intestine and also prevents backflow of faeces into the ileum.
The microscopic anatomy of the small intestine is designed for maximal nutrient absorption. It contains circular folds, villi and microvilli, which along with its length, create a huge surface area. The circular folds found in the mucosa and submucosa force chyme slowly through the lumen, allowing time for absorption of nutrients. Fingerlike projections of the mucosa, called villi, are large in duodenum and gradually get smaller and shorter as progress is made through the small intestine. Villi are composed of absorptive, columnar, epithelial cells, known as enterocytes, which are held together by tight junctions. Mucous-secreting goblet cells are found interspersed between these epithelial cells. Between the villi the mucosa is dotted with pits that lead to tubular glands known as intestinal crypts. Crypt epithelial cells secrete intestinal juice, while paneth cells located at the base of each crypt secrete lysozyme and defensin proteins, both of which protect the mucosa from bacterial invasion and help control the microflora (normal flora) of the small intestine. About 1-2 L of intestinal juice is secreted by crypt cells daily in response to the acid in the chyme and distension of the small intestine as it enters the duodenum. Intestinal juice contains water and mucous, but very little enzyme. Enzymes active in the small intestine come mainly from the brush border of the small intestine and pancreatic juice. These enzymes work best in an alkaline environment.
Laying amongst the crypt cells are enteroendocrine cells which secrete the hormones secretin and cholecystokinin (CCK). The core of each villus contains an arteriole, blood capillaries, a venule and a lymphatic capillary known as a lacteal. The blood capillaries absorb most nutrients with the lacteal absorbing most lipids. On the surface of the villi are microvilli which form a brush border (so named because they look like the strands of a brush under the light microscope). The plasma membrane of the microvilli contains brush border enzymes which complete the digestion of carbohydrates and proteins in the small intestine.
The ileum lies within the hypogastric region and has a thinner, less muscular and less vascular wall. Lymphatic nodules known as Peyer patches are found attached to the ileum, becoming larger as the ileum approaches the large intestine. The ileum joins the cecum of the large intestine at the ileocecal junction, where a muscular sphincter, the ileocecal valve, is found. This valve controls the passage of food into the large intestine and also prevents backflow of faeces into the ileum.
The microscopic anatomy of the small intestine is designed for maximal nutrient absorption. It contains circular folds, villi and microvilli, which along with its length, create a huge surface area. The circular folds found in the mucosa and submucosa force chyme slowly through the lumen, allowing time for absorption of nutrients. Fingerlike projections of the mucosa, called villi, are large in duodenum and gradually get smaller and shorter as progress is made through the small intestine. Villi are composed of absorptive, columnar, epithelial cells, known as enterocytes, which are held together by tight junctions. Mucous-secreting goblet cells are found interspersed between these epithelial cells. Between the villi the mucosa is dotted with pits that lead to tubular glands known as intestinal crypts. Crypt epithelial cells secrete intestinal juice, while paneth cells located at the base of each crypt secrete lysozyme and defensin proteins, both of which protect the mucosa from bacterial invasion and help control the microflora (normal flora) of the small intestine. About 1-2 L of intestinal juice is secreted by crypt cells daily in response to the acid in the chyme and distension of the small intestine as it enters the duodenum. Intestinal juice contains water and mucous, but very little enzyme. Enzymes active in the small intestine come mainly from the brush border of the small intestine and pancreatic juice. These enzymes work best in an alkaline environment.
Laying amongst the crypt cells are enteroendocrine cells which secrete the hormones secretin and cholecystokinin (CCK). The core of each villus contains an arteriole, blood capillaries, a venule and a lymphatic capillary known as a lacteal. The blood capillaries absorb most nutrients with the lacteal absorbing most lipids. On the surface of the villi are microvilli which form a brush border (so named because they look like the strands of a brush under the light microscope). The plasma membrane of the microvilli contains brush border enzymes which complete the digestion of carbohydrates and proteins in the small intestine.
Accessory organs of the GIT
The liver, gall bladder and pancreas all have digestive functions associated with the small intestine.
The liver is the largest organ in the human body and has numerous metabolic functions which include:
The liver's primary digestive function is to produce bile which is transported to the gall bladder where it is stored until needed.
Bile travels from the gall bladder via the bile duct to be released into the duodenum along with pancreatic juice from the pancreas. It is a yellow-green alkaline solution containing bile salts which emulsify fats; that is, it breaks them down into fats droplets increasing the surface area of the fats so that fat-digesting enzymes can act on the lipids. Bile salts also aid in the absorption of fats and cholesterol.
The pancreas is both an accessory digestive organ and an endocrine gland. For digestion, it produces pancreatic enzymes that breakdown a variety of food products; proteins, lipids and nucleic acids. Pancreatic juice, which enters the duodenum via the pancreatic duct contains various enzymes, water and bicarbonate ions. About 1.2-1.5 L is produced daily and its alkaline pH helps to neutralise the acidic chyme leaving the stomach. The endocrine function of the pancreas is related to blood glucose maintainence which is discussed later under the heading ‘blood glucose homeostasis’
The liver is the largest organ in the human body and has numerous metabolic functions which include:
- Processing nutrients carried in the blood,
- Storage of glucose as glycogen,
- Storage of fat soluble vitamins,
- Detoxification.
The liver's primary digestive function is to produce bile which is transported to the gall bladder where it is stored until needed.
Bile travels from the gall bladder via the bile duct to be released into the duodenum along with pancreatic juice from the pancreas. It is a yellow-green alkaline solution containing bile salts which emulsify fats; that is, it breaks them down into fats droplets increasing the surface area of the fats so that fat-digesting enzymes can act on the lipids. Bile salts also aid in the absorption of fats and cholesterol.
The pancreas is both an accessory digestive organ and an endocrine gland. For digestion, it produces pancreatic enzymes that breakdown a variety of food products; proteins, lipids and nucleic acids. Pancreatic juice, which enters the duodenum via the pancreatic duct contains various enzymes, water and bicarbonate ions. About 1.2-1.5 L is produced daily and its alkaline pH helps to neutralise the acidic chyme leaving the stomach. The endocrine function of the pancreas is related to blood glucose maintainence which is discussed later under the heading ‘blood glucose homeostasis’
Digestion and absorption in the small intestine: carbohydrates
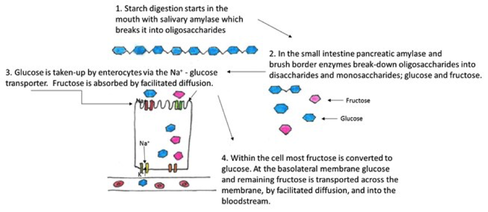
Digestion and absorption of starch (adapted from Saladin, 2015. & Mckinley, 2015).
About 50 % of starch (polysaccharides) in the food has been digested by the time it reaches the small intestine. Once in the duodenum the release of pancreatic amylase resumes the process. Within 10 min of entering the small intestine starch has been broken-down into oligosaccharides, mainly maltose. These oligosaccharides are further digested by brush border enzymes. Eventually all the starch will be reduced to monosaccharides.
About 80 % of absorbed monosaccharides are glucose molecules that are moved from the lumen of the small intestine into the epithelial cells by the sodium-glucose transporter. Glucose then exits the basolateral membrane of the cell by facilitated diffusion and enters the blood capillaries through intercellular clefts
About 80 % of absorbed monosaccharides are glucose molecules that are moved from the lumen of the small intestine into the epithelial cells by the sodium-glucose transporter. Glucose then exits the basolateral membrane of the cell by facilitated diffusion and enters the blood capillaries through intercellular clefts
Digestion and absorption in the small intestine: proteins
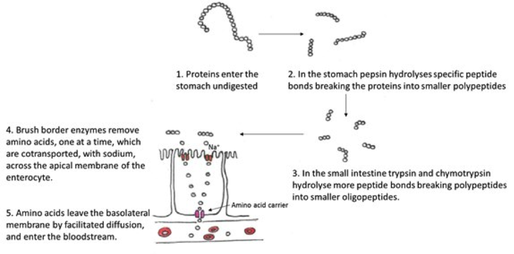
Digestion and absorption of proteins (adapted from Saladin, 2015. & Mckinley, 2015).
Whole proteins are not usually absorbed. It is the protein constituents, the amino acids, that come from dietary proteins, digested enzymes (enzymes are proteins in structure), and digested epithelial cells that are absorbed into the blood stream. Enzymes known as proteases, in particular pepsin, start the initial break-down of proteins in the stomach. Shorter peptides and some amino acids move into the small intestine where trypsin (from the pancreas) and chymotrypsin continue protein digestion into to oligopeptides. Finally enzymes break the oligopeptides into individual amino acids. The breakdown sequence is
Whole proteins -> polypeptides -> oligopeptides -> amino acids
Absorption of amino acids is like that of glucose. Enterocytes have several types of carriers to transport the different amino acids, usually coupled to the active transport of sodium. Dipeptides and tripeptides are actively absorbed by H+ -dependent cotransport where they are broken-down into their individual amino acids within the epithelial cell. Entry into the capillary is by diffusion.
Whole proteins -> polypeptides -> oligopeptides -> amino acids
Absorption of amino acids is like that of glucose. Enterocytes have several types of carriers to transport the different amino acids, usually coupled to the active transport of sodium. Dipeptides and tripeptides are actively absorbed by H+ -dependent cotransport where they are broken-down into their individual amino acids within the epithelial cell. Entry into the capillary is by diffusion.
Digestion and absorption in the small intestine: lipids
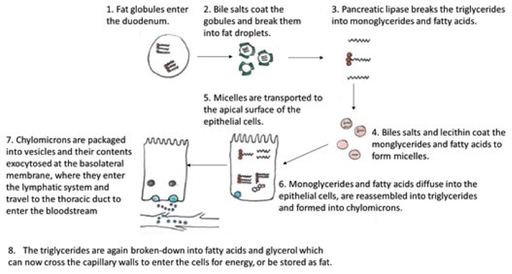
Digestion and absorption of lipids (adapted from Saladin, 2015. & Mckinley, 2015).
Fats are digested by enzymes called lipases. Starting in the mouth lingual lipase in saliva begins to digest a small amount of any fat in the food. Once in the acidic environment of the stomach along with gastric lipase the digestive process continues. The churning movements of the stomach serves to break the fats into small droplets dispersed within the chyme which then enters the duodenum. Segmentation within the duodenum breaks-down the fat even further, and the components of bile, coat these small droplets of fat within the duodenum, preventing them from reforming into larger globules. Pancreatic lipase further contributes to fatty acid break-down by removing the first and third fatty acid from triglycerides, resulting in a monoglyceride and two fatty acid chains.
Absorption of fats depends on the action of tiny droplets in the bile, micelles, into which bile phospholipids and cholesterol diffuse to form a micelle core. Within the duodenum fat soluble vitamins, cholesterol, fatty acid chains and monoglycerides are absorbed into the micelles which transport the lipids to the surface of enterocytes where they leave the micelles and diffuse across the plasma membrane into the cell. Within the cell the fatty acids and monoglycerides are reconstituted to make triglycerides which are in turn coated with phospholipids and cholesterol to form water-soluble chylomicrons. These chylomicrons are packaged into vesicles which move to the basolateral membrane where they are exocytosed and enter lacteals of the lymphatic system. Fat travels in lymph to the thoracic duct and enters the bloodstream at the left subclavian vein. Once in the bloodstream lipoprotein lipase hydrolyses the triglycerides to free fatty acids and glycerol, which can then pass across the capillary wall to be used for energy or stored as fat.
Absorption of fats depends on the action of tiny droplets in the bile, micelles, into which bile phospholipids and cholesterol diffuse to form a micelle core. Within the duodenum fat soluble vitamins, cholesterol, fatty acid chains and monoglycerides are absorbed into the micelles which transport the lipids to the surface of enterocytes where they leave the micelles and diffuse across the plasma membrane into the cell. Within the cell the fatty acids and monoglycerides are reconstituted to make triglycerides which are in turn coated with phospholipids and cholesterol to form water-soluble chylomicrons. These chylomicrons are packaged into vesicles which move to the basolateral membrane where they are exocytosed and enter lacteals of the lymphatic system. Fat travels in lymph to the thoracic duct and enters the bloodstream at the left subclavian vein. Once in the bloodstream lipoprotein lipase hydrolyses the triglycerides to free fatty acids and glycerol, which can then pass across the capillary wall to be used for energy or stored as fat.
Digestion and absorption in the large intestine
The large intestine is divided into the cecum, appendix, colon, rectum and anal canal. The cecum is the first part of the large intestine and joins the ileum at the ileocecal valve. The appendix is a mass of lymphoid tissue attached to the posteromedial surface of the cecum, and has a role in immunity. The colon is divide in to the ascending, transverse and descending regions, which on entering the pelvis becomes the S-shaped sigmoid colon.
Anatomically the large intestine differs from the small intestine, its mucosa being simple columnar epithelium. There are no circular folds, or villi present, and cells that produce digestive enzymes are few. Why do you think this is? The walls of the large intestine are thicker than that of the small intestine, with deeper crypts that contain numerous mucous-secreting cells. The anal canal differs from the rest of the large intestine in that its mucosa is stratified squamous epithelium, the superior surface of which hangs in long ridges called anal columns. Between the anal columns are anal sinuses which produce mucous when compressed, which aids the emptying of the bowel.
Within the large intestine are numerous bacteria which play an important role in digestion. They;
Most digestive processes are complete by the time the chyme reaches the large intestine and no further break-down of substances occurs here.
As you can see at each stage of digestion different enzymes are released to breakdown specific nutrients into smaller components.
The table below summarises the various parts of the system and the processes that take place along the digestive tract.
Anatomically the large intestine differs from the small intestine, its mucosa being simple columnar epithelium. There are no circular folds, or villi present, and cells that produce digestive enzymes are few. Why do you think this is? The walls of the large intestine are thicker than that of the small intestine, with deeper crypts that contain numerous mucous-secreting cells. The anal canal differs from the rest of the large intestine in that its mucosa is stratified squamous epithelium, the superior surface of which hangs in long ridges called anal columns. Between the anal columns are anal sinuses which produce mucous when compressed, which aids the emptying of the bowel.
Within the large intestine are numerous bacteria which play an important role in digestion. They;
- Synthesise B complex vitamins
- Synthesise some vitamin K
- Metabolise mucin, heparin and hyaluronic acid
- Ferment indigestible carbohydrates.
Most digestive processes are complete by the time the chyme reaches the large intestine and no further break-down of substances occurs here.
As you can see at each stage of digestion different enzymes are released to breakdown specific nutrients into smaller components.
The table below summarises the various parts of the system and the processes that take place along the digestive tract.

Summary of digestive processes throughout the GIT
4 Metabolism
Once nutrients have been digested and absorbed they are involved in a myriad of biochemical processes known as metabolism.
These metabolic processes either build new materials, termed anabolism, or breakdown complex structures to simpler ones, termed catabolism.
Catabolic reactions use nutrients, in particular glucose, to create energy in the form of ATP. This process is known as cellular respiration. ATP provides the energy to drive cellular work. For ingested food to provide energy it has to first be digested, absorbed and carried to the cells. Within the cytoplasm of the cells the nutrients are either anabolised to from proteins, lipids and glycogen, or catabolised to form pyruvic acid. Finally, within the cell’s mitochondria, in the presence of oxygen, breakdown is completed with large quantities of ATP being produced.
Carbohydrates are ultimately broken-down into the monosaccharide glucose. There are three major pathways for glucose catabolism:
Lipid catabolism begins with the hydrolysis of triglycerides into glycerol and fatty acids. Glycerol is converted to glyceraldehyde 3-phosphate (PGAL), and enters the glycolytic pathway and yields 18 ATP. Fatty acids are catabolised in the mitochondria by a process known as beta-oxidation, the products of which is acetyl groups, which combine with co-enzyme A, to produce acetyl-CoA. Acetyl CoA then enters the citric acids cycle. By this pathway, a 16-carbon fatty acid can yield 129 ATP molecules.
Proteins are broken-down into free amino acids which can be used along with dietary amino acids to form new proteins. Free amino acids can also be converted to glucose by a three-step process; deamination, amination and transamination. The liver is responsible for nutrient interconversion.
These metabolic processes either build new materials, termed anabolism, or breakdown complex structures to simpler ones, termed catabolism.
Catabolic reactions use nutrients, in particular glucose, to create energy in the form of ATP. This process is known as cellular respiration. ATP provides the energy to drive cellular work. For ingested food to provide energy it has to first be digested, absorbed and carried to the cells. Within the cytoplasm of the cells the nutrients are either anabolised to from proteins, lipids and glycogen, or catabolised to form pyruvic acid. Finally, within the cell’s mitochondria, in the presence of oxygen, breakdown is completed with large quantities of ATP being produced.
Carbohydrates are ultimately broken-down into the monosaccharide glucose. There are three major pathways for glucose catabolism:
- Glycolysis - yields two pyruvic acid molecules from each glucose molecule. 2 ATP molecules are produced.
- Anaerobic fermentation - reduces the pyruvic acid to lactic acid. No ATP is produced, but this process allows glycolysis to continue in the absence of oxygen.
- Aerobic respiration – oxidation of pyruvic acid to carbon dioxide and water in the presence of oxygen. 36 ATP molecules are produced in a series of reactions known as the citric acid or Krebs cycle.
Lipid catabolism begins with the hydrolysis of triglycerides into glycerol and fatty acids. Glycerol is converted to glyceraldehyde 3-phosphate (PGAL), and enters the glycolytic pathway and yields 18 ATP. Fatty acids are catabolised in the mitochondria by a process known as beta-oxidation, the products of which is acetyl groups, which combine with co-enzyme A, to produce acetyl-CoA. Acetyl CoA then enters the citric acids cycle. By this pathway, a 16-carbon fatty acid can yield 129 ATP molecules.
Proteins are broken-down into free amino acids which can be used along with dietary amino acids to form new proteins. Free amino acids can also be converted to glucose by a three-step process; deamination, amination and transamination. The liver is responsible for nutrient interconversion.
5 Metabolic states
Throughout a 24-hour period metabolism varies depending on whether one has eaten, of if you are between meals or you are, for example exercising. This variation is referred to as the metabolic rate. It is measured as the amount of energy released in the body per unit of time. There is a minimum amount of energy required for the body to maintain all its functions and is known as the basal metabolic rate (BMR). This is measured between 12-14 hours after a meal, when one is awake, relaxed and at a comfortable temperature. The total metabolic rate (TMR) is the sum of the BMR plus any additional energy used due to physical and mental activity. Energy consumed should equal energy output, and when this is the case body weight remains stable, an imbalance leads to either weight gain or weight loss.
Absorbed nutrients are continuously being catabolised and anabolised depending on energy requirements and the time since the last meal. Metabolic controls maintain blood nutrient levels between two states; the absorptive state and the post-absorptive state. The absorptive state is time following a meal and lasts for about four hours. During this time food is being digested, moves into the blood stream and is converted into energy stores. Glucose is readily available to synthesise ATP, and so anabolism exceeds catabolism. During the post-absorptive state the GIT is empty and so catabolism exceeds anabolism. The body must draw on stored fuels to maintain blood glucose levels for the production of ATP.
Absorbed nutrients are continuously being catabolised and anabolised depending on energy requirements and the time since the last meal. Metabolic controls maintain blood nutrient levels between two states; the absorptive state and the post-absorptive state. The absorptive state is time following a meal and lasts for about four hours. During this time food is being digested, moves into the blood stream and is converted into energy stores. Glucose is readily available to synthesise ATP, and so anabolism exceeds catabolism. During the post-absorptive state the GIT is empty and so catabolism exceeds anabolism. The body must draw on stored fuels to maintain blood glucose levels for the production of ATP.
Blood glucose homeostasis
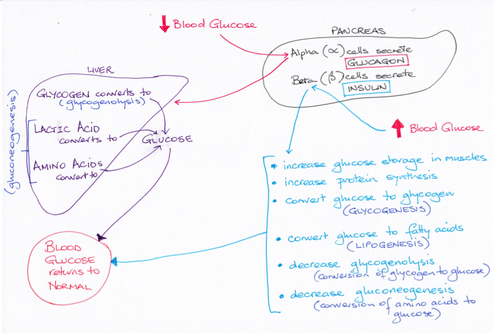
Glucose levels within the blood are kept within tight parameters between 4.0–7.8 mmol/L. To achieve this, two hormones are secreted by the pancreas in response to rising and falling glucose levels. In the absorptive state glucose levels rise rapidly, and along with the intestinal hormones secretin, gastrin and CCK, trigger the beta cells of the pancreatic islets to secrete insulin. Insulin binds to receptors on the membrane of target cells and cellular uptake of glucose increases. Insulin also stimulates glucose oxidation, glycogenesis and lipogenesis, inhibits gluconeogenesis. The uptake amino acids into cells and subsequent protein synthesis is increased. The result of insulin’s actions leads to falling blood glucose levels.
In the post-absorptive state levels of insulin decline as blood glucose levels fall. The lower levels of circulating glucose stimulate the alpha cells of the pancreatic islets to secrete the hormone glucagon. Glucagon acts on the liver, stimulating glycogenolysis and gluconeogenesis, while adipose tissue undergoes lipolysis releasing fatty acids and glycerol into the blood. Thus, blood glucose levels start rise.
In the post-absorptive state levels of insulin decline as blood glucose levels fall. The lower levels of circulating glucose stimulate the alpha cells of the pancreatic islets to secrete the hormone glucagon. Glucagon acts on the liver, stimulating glycogenolysis and gluconeogenesis, while adipose tissue undergoes lipolysis releasing fatty acids and glycerol into the blood. Thus, blood glucose levels start rise.
Temperature regulation
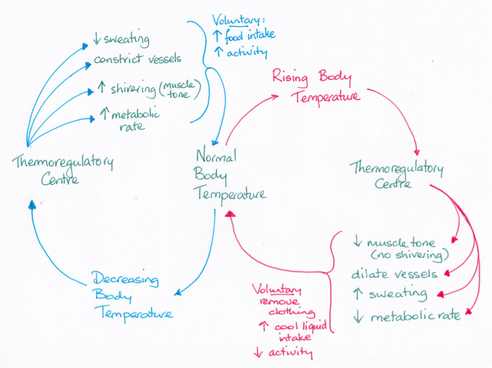
Temperature Regulation
Heat is produced from the metabolic reactions that take place within the body. At rest, most heat is produced by the heart, brain, liver and endocrine organs. While skeletal muscles generate about 25 % of heat when at rest, during exercise they can produce up to 40 times more heat than the rest of the body. While heat production can vary depending on activity level, core body temperature is maintained within tight parameters between 37.2 degrees Celsius and 37.6 degrees Celsius and does not vary by more than about 1 degree Celsius in any 24-hour period. The core temperature is important for maintaining the internal organs at an operant temperature. The shell temperature, at the skin surface and the mouth, is where heat is lost and can vary considerably.
6 Important roles of the liver
Aside from metabolic and digestive functions, the liver detoxifies alcohol, antibiotics and other drugs, metabolises bilirubin from red blood cell break-down, and excretes bile pigments. The liver contains macrophages, known as Kupffer cells, which remove bacteria and foreign materials from the blood. The liver also plays a role in steroid hormone removal from the body preventing them reaching excess levels. You can see why disease of the liver can have far-reaching effects on the body.
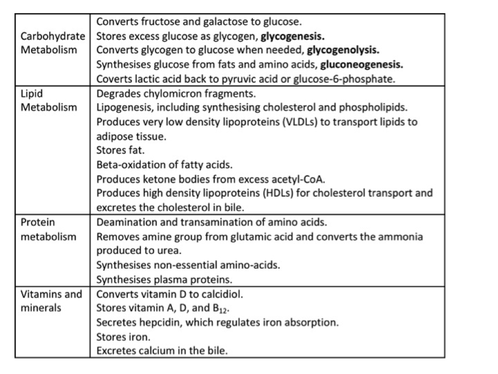
Major functions of the liver hepatocytes
References
Marieb, E. N. & Hoehn,K. (2013). Human Anatomy and Physiology. Pearson Education, USA. (9th Ed).
Mckinley, M. P., O’Loughlin, V. D. & Bidle, T. S. (2015). Anatomy and Physiology; An Integrative Approach.
Saladin. K. S. (2015). Anatomy and Physiology; The Unity of Form and Function. McGraw-Hill Education, USA. (7th Ed).
Mckinley, M. P., O’Loughlin, V. D. & Bidle, T. S. (2015). Anatomy and Physiology; An Integrative Approach.
Saladin. K. S. (2015). Anatomy and Physiology; The Unity of Form and Function. McGraw-Hill Education, USA. (7th Ed).

